![]()
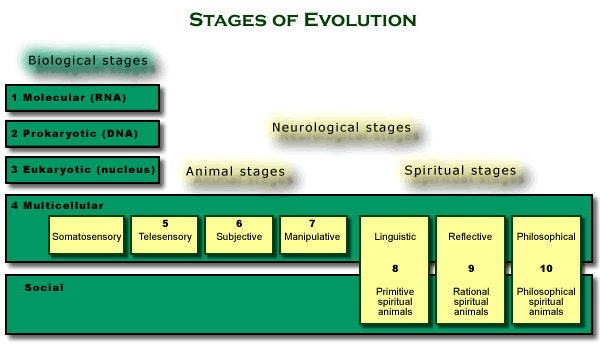
 8 Linguistic stage (primitive spiritual animals). We
have followed the series of levels of neurological organization, tracing how
multicellular animals evolve, stage by stage, from somatosensory animals,
through telesensory and subjective animals, to manipulative animals. In each
case, the increase in animal power came from increases in the capacity of
the animal system of representation to internalize spatial aspects
of the world. We now take up the final three stages of evolution leading up
to beings like us. Though they continue the series of stages caused by levels
of neurological organization, they also involve something quite novel. Each
of these last three stages is also caused by a higher level of neurological organization,
but they are levels of organization in a linguistic system of
representation, and the function of language is not just how it increases
the power of individual animals, but also how it increases the power of groups
of individuals. What is novel about these stages is that they are also the
evolution of spiritual animals.
8 Linguistic stage (primitive spiritual animals). We
have followed the series of levels of neurological organization, tracing how
multicellular animals evolve, stage by stage, from somatosensory animals,
through telesensory and subjective animals, to manipulative animals. In each
case, the increase in animal power came from increases in the capacity of
the animal system of representation to internalize spatial aspects
of the world. We now take up the final three stages of evolution leading up
to beings like us. Though they continue the series of stages caused by levels
of neurological organization, they also involve something quite novel. Each
of these last three stages is also caused by a higher level of neurological organization,
but they are levels of organization in a linguistic system of
representation, and the function of language is not just how it increases
the power of individual animals, but also how it increases the power of groups
of individuals. What is novel about these stages is that they are also the
evolution of spiritual animals.
The linguistic system of representation is built on the animal system of representation, but it does not increase the power of individual animals, at least, not originally. Just as higher levels of neurological organization introduced, first, spatial imagination and, then, structural imagination, to the animal system of representation, so each of these three yet higher levels of neurological organization introduces new forms of imagination in the linguistic system of representation (naturalistic, rational and philosophical imagination, as we shall see). And just as the new forms of imagination in the animal system of representation made individual animals more powerful by enabling them to adapt their behavior to spatial aspects of the world, so these new forms of imagination in the linguistic system of representation do eventually make individual animals more powerful by enabling them to adapt their behavior to further aspects of the world. However, such contributions to the power of individual animals are, at most, only half of the story. (The new forms of imagination in the linguistic system of representation are the blue panels in the accompanying diagram.)
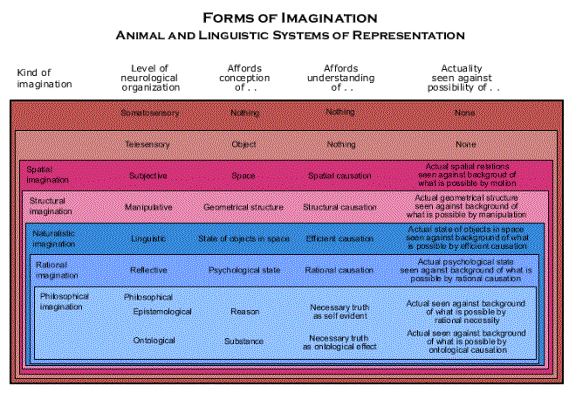
The linguistic system of representation is not just a new kind of imagination in the individual, but also a new form of interaction among individuals. Linguistic interactions among individuals make groups of animals more powerful as a whole, and that is the original function of language, from which the increase in individual power derives. Indeed, it is a source such great power that groups of language using animals constitute animals on the social level of biological organization. It is, as we shall see, a new form of life, and such groups will be called “spiritual animals.”
What makes these final three stages different from all earlier stages is that organisms are evolving on two levels of biological organization at once. Not only do individual members, by their reproduction, impose natural selection on themselves at the individual level, but the social level animals they constitute impose natural selection on themselves at the social level by their reproduction at the social level. Thus, gradual evolution during these final three stages is change in the direction of the natural perfection of organisms at both levels of biological organization at once. And since they evolve together, there is also a natural perfection about their combination.
To mark this difference, these final three stages will be called “spiritual stages of evolution,” distinguishing them from the multicellular animal stages of evolution just discussed, and the animals evolving on the social level will be called “spiritual animals.” The appropriateness of calling the “spiritual” will be explained after we explain the function of language.
Since these final three levels of neurological organization are also stages in the evolution of a new kind of organism on the social level, the first stage has two ontological causes at once, according to this ontological explanation of evolutionary stages as a revolutionary global regularity, both a higher level of neurological organization and a higher level of biological organization. Once spiritual animals have evolved, subsequent evolutionary stages are caused ontologically by higher levels of neurological evolution in the individual multicellular animals, though their higher levels of organization are also evident in the linguistic representation they exchange.
Primitive spiritual animals. Primitive spiritual animals are groups of primates with the capacity to use a language of natural sentences (that is, sentences with a simple subject-predicate grammar). The use of language is functional, as we shall see, because it enables their behavior to be coordinated so that they can act as a whole. It is a new animal behavior guidance system that is possible only at the social level of biological organization. But language requires a higher level of part-whole complexity, the linguistic level of neurological organization, and such a higher level is possible, as we shall see, because it evolves within the faculty of imagination. The new form of imagination, called “naturalistic imagination,” gives subjective animals the conception of a state of affairs in the natural world, and that enables them to understand efficient causation.
Rational spiritual animals. Rational spiritual animals are also animals at the social level of biological organization, but they have the capacity to use psychological sentences (that is, sentences with predicates formed of verbs of propositional attitude, such as “believes” and “desires,” together with complete sentences). We shall see what its original function was, and how it too was possible because of a higher level of neurological organization in the faculty of imagination. It is aptly called the reflective level of neurological organization, because this higher level brain gives subjective animals the conception of a psychological state and enables them to think about such states as causes of behavior (and belief). The new form of imagination it gives them will be called “rational imagination,” because as they use psychological sentences to think about the causes of their own behavior, the causes of their behavior become reasons. (Reasons are causes of behavior that are represented as causing behavior as part of the very process of causing behavior.) Rational imagination gives them an understanding of rational causation, but since that enables them to understand arguments, it is also the foundation for a new form of evolution by reproductive causation, namely, cultural evolution, which is contained within spiritual animals. Thus, reason makes language a very powerful animal behavior guidance system, which takes over responsibility for behavior at both the individual and social level.
Philosophical spiritual animals. Philosophical spiritual animals are social level animals in which the arguments accumulated as culture have evolved a higher level of organization. That is, in philosophical arguments, all the arguments of the rational spiritual level are organized as parts of a single argument. That promises a new way of proving that propositions are true, and thus, it is a new way of discovering the true and the good, one that yields necessarily truths in one way or another. Though this is a stage of cultural evolution, it gives the members of the spiritual animals in which it exists a higher level of rational imagination, and thus, it is also the philosophical level of neurological organization. But there are, as we have seen, two ways of constructing such philosophical arguments, epistemological and ontological, and only one of them is able to succeed in the end.
Primitive spiritual animals evolve at the first spiritual stage of evolution. In order to show that a stage of evolution is inevitable, according to reproductive causation, it is necessary to show that the higher level of part-whole complexity in the structure of evolving organisms is both functional and possible.
The higher level must be functional in the sense of opening up an entire new range of powers for controlling relevant conditions, which can evolve gradually over the stage, and it must be possible in the sense that the higher level of organization can be tried out as a random variation in the evolving organisms.
In the case of primitive spiritual animals, however, there are two kinds of higher levels of part-whole complexity, a higher neurological level and a higher biological level, and the simplest way to explain their function is to assume, for now, that the linguistic level of neurological organization is possible, and to explain its origin by its functions, that is, by how it was naturally selected.
That will put us in a position to consider the nature of spiritual animals in the second section.
Only in the third section will I take up the issue about the possibility of the linguistic level of neurological organization, and that will include an explanation of the structure of the linguistic brain and how naturalistic imagination affords an understanding of efficient causation.
Finally, in the fourth section, I will consider the empirical evidence about the gradual evolution of primitive spiritual animals on earth.
 Function
of the linguistic level of neurological organization. The original function of a level of neurological
organization higher than manipulative animals is a power it gave to entire
groups of such animals, and thus, the easiest way to explain its function
is to consider how spiritual animals evolved from animal societies. Thus,
we begin with the radiation of primates. Such a radiation was inevitable,
once higher primates had evolved, and it would lead some higher primates out
of the forest, if it was possible, because that would open up new sources
of free energy.
Function
of the linguistic level of neurological organization. The original function of a level of neurological
organization higher than manipulative animals is a power it gave to entire
groups of such animals, and thus, the easiest way to explain its function
is to consider how spiritual animals evolved from animal societies. Thus,
we begin with the radiation of primates. Such a radiation was inevitable,
once higher primates had evolved, and it would lead some higher primates out
of the forest, if it was possible, because that would open up new sources
of free energy.
With the addition of structural imagination, higher primates were more powerful than other mammals, and though they started off simple, uniform and weak, manipulative animals gradually became more complex, diverse, and powerful. Such gradual evolution is change in the direction of maximum holistic power at both the level of the organisms and the ecology, which is natural perfection for organisms of their kind. Thus, as each primate species became as powerful as possible at controlling relevant conditions in its ecological niche, new primate species were added, dividing up the sources of usable energy and invading new habitats, and species generally adapted to one another, maximizing the use of the available free energy to fuel reproductive cycles generally. It was inevitable, because, as we have seen, reproductive causation always makes every possible increase in power actual.
 Hominids.
The radiation of primates beyond their original arboreal habitat was just
a special case of species tapping new sources of free energy. As primates
diversified to fill all possible ecological niches, some invaded the grasslands
beyond the forest to tap the energy available there (such as grass seeds,
berries and small animals). These primates evolved into hominids as their
bodies and behavior became as powerful as possible in controlling the conditions
that affected their reproduction in this new ecological niche. The traits
that primates would evolve, once they invaded the grasslands, are predicable,
in principle, because only a certain range of random variations can be tried
out by their biological behavior guidance system and the conditions affecting
their reproduction were fixed by their new ecological niche.
Hominids.
The radiation of primates beyond their original arboreal habitat was just
a special case of species tapping new sources of free energy. As primates
diversified to fill all possible ecological niches, some invaded the grasslands
beyond the forest to tap the energy available there (such as grass seeds,
berries and small animals). These primates evolved into hominids as their
bodies and behavior became as powerful as possible in controlling the conditions
that affected their reproduction in this new ecological niche. The traits
that primates would evolve, once they invaded the grasslands, are predicable,
in principle, because only a certain range of random variations can be tried
out by their biological behavior guidance system and the conditions affecting
their reproduction were fixed by their new ecological niche.
Carrying clubs. The main obstacle to occupying the grasslands were carnivores, such as lions and packs of wolves. The ungulates already inhabiting the grasslands protected themselves by traveling in herds and running away from these predators. But primates are relatively slow-moving, which made them easy prey. However, grasslands were not closed to them, because, as manipulative animals, they had another means of protection, not available to other mammals. They could carry clubs into the grassland with them and beat off carnivores that tried to prey on them.
Social life. This means of protection would be effective, however, only if several adults traveled together carrying clubs. Predators might still overcome one or two primates with clubs, especially if they were parents with children to protect. However, several adult primates carrying clubs could attack simultaneously from different directions, and that would thwart carnivores that were adapted to preying on ungulates, including not only the most ferocious, but also packs of smaller predators.
The need to stay together in a group would not be a great obstacle to their evolution, because group life in the forest had already given higher primates desires that enabled multiple families to live together in groups. They had instinctive desires that would establish a dominance hierarchy among them, and conflicts would be resolved by some members having power over others. Thus, grassland energy sources were open to any small group of primates that carried clubs and had learned how to wield them. This variation was well within the range of those being tried out randomly by existing primates, especially considering that, at first, nomadic primates could return to the forest at night for protection.
Since neither club-carrying nor social life was optional for nomadic primates, the evolution of two kinds of traits were inevitable, one a change in the biological behavior guidance system and the other a change mainly in the animal behavior guidance system set up by it.
Bipedal stance. The need to carry clubs entails the evolution of a bipedal stance in the primates that invaded the grassland. On the ground, primates were naturally disposed, like most mammals, to travel on all fours. And the further they could travel in gathering food, the wider the range of grasslands that they could invade. But nomadic primates could not travel on all fours, if they had to carry clubs along with them. As manipulative animals, however, they had another alternative. The precise control of muscles in their hands enabled primates to balance their bodies on two of their hands, leaving the other two hands free to carry clubs. Thus, the primates that invaded the grasslands would eventually acquire bodies suited to bipedal locomotion: random variations in embryological development that made them better able to balance their bodies on two legs controlled a condition that affected their reproduction and, thus, was selected as part of their biological behavior guidance systems. The primate predecessors of humans who walked upright, on two feet, are now generally known as “hominids”.
Although the traditional evolutionary explanation of bipedalism is also that it evolved for carrying things, this explanation is different. A century ago, Darwinians explained bipedalism by the need to carry tools. But that did not really explain bipedalism, because they attributed tool use to an increase in brain size or intelligence without being able to explain why an increase in brain size was necessary.
Sociobiologists have now dropped the assumption that brain size naturally increases, but their explanation does not make increasing brain size any more necessary. They claim that bipedalism evolved in order to carry the food required for the practice of food sharing. But that is no explanation, for they do not explain why hominids must share food. Apparently, increasing sociality has replaced increasing intelligence as the motor behind hominid evolution. (See, for example, Wilson and Lumsden 1983, pp. 10-12.)
To insist that the cause of bipedalism was the need to carry clubs for protection is not to deny that hominids also carried other tools. Nor to deny that they shared food. It is only to insist that bipedalism be explained by the function that made the trait inevitable, that is, by the one that predicts its evolution, rather than other functions which were made possible by its evolution. There is no reason to believe that bipedalism would have evolved to carry other tools or to share food, if nomadic primates did not have to carry clubs. And it would have evolved to carry clubs, even if it did not have those other functions.
Altruism. Another set of traits can be predicted by another function that needed to be served in their ecological niche. Protection from predators required not only traveling in groups, but also a disposition to use their clubs to assist other members of the group when they were endangered. Since this exposes the protectors to greater danger than sitting out the attack or running away, at least, in the short run, it is an example of altruism.
Altruism poses a challenge to explaining the evolution of nomadic bands of primates, therefore, because it seems that altruism cannot evolve by the natural selection of reproducing organisms. If individual animals are naturally selected by their success in reproducing, traits that benefit others at the expense of the animals conferring the benefits, then natural selection will eliminate individuals with those traits in favor of those who receive the benefits.
The altruism required of nomadic hominids resembles the bees’ use of stingers to protect the hive, but bees cannot be the model for explaining primate altruism. Defender bees die as they use their stingers to protect the hive, and that can be explained in social insects, because individual bees are parts of a multisomatic organism set up by way of a single fertilized egg cell. By including some members with stingers, the insect colony is able to do the non-reproductive work of protecting the hive as a whole. But hominid groups are not multisomatic organisms, because their member continue to reproduce sexually as individuals. Thus, another explanation is needed for the trait of protecting other members of their nomadic group.
The received solution to this puzzle about altruism is called “kin selection,” but it depends on a theory of evolution, called the “selfish gene,” which is incompatible with reproductive causation. The received view is, however, pointing to a relevant cause, which is explained in another way by reproductive causation. Though it does not quite explain the phenomenon of altruism in hominids, it will serve to introduce a cause that does make altruism inevitable.
What makes it appear that the gene is the unit of selection is, as we have seen, the sexual mixing of lower level structural causes in the process of reproduction, for it means that as organisms are evolving greater power, natural selection is focused on only certain of their genes (or chromosomes), which distinguish individuals from one another. Sexual reproduction is a way of internalizing reproductive causation so that lower level structural causes within the organism continue to evolve as the organism goes through reproductive cycles. And thus gradual evolution can be seen as genes competing with one another to appear in the next generation by the traits that are their non-reproductive structural effects, that is, as “selfish genes.”
The selfish gene theory explains altruistic behavior by the fact that siblings are likely to share the same genes. Thus, if altruistic behavior increases the likelihood of their siblings succeeding in reproducing more than it costs in terms of the animals own likelihood of reproducing, then the gene will be selected to appear in the next generation.
What the theory of kin selection of selfish genes is pointing to is another aspect of the evolution of the sexual mixing of genetic structures in reproduction. The obstacle to explaining the evolution of altruism is only an appearance that comes from failing to recognize that what is actually going through reproductive cycles, the primary structure, is not the individual multicellular animal, but the mating pair — or even the whole family.
The basic ontological cause of gradual evolution is, as we have seen, the reproductive cycle, because reproduction is what combines with space to make free energy scarce and that is how material structures that generate the whole cycles, including both reproductive and non-reproductive work, impose natural selection on themselves. But in the case of sexually reproducing animals, as we saw when the origin of sex was explained in eukaryotes (Stage 3), the individual animal is not that whole primary structure, because it cannot reproduce by itself. The individual organism must mate in order to reproduce, and that means, in effect, finding its other half. The mating pair is, therefore, the primary structure which is shaped by reproductive causation.
The primary structure is the “unit of selection,” and so the unit of selection is larger than the individual (rather than smaller, as the selfish gene hypothesis would have it), for it includes both members of the mating pair (though the pair that makes up a particular reproducing organism is not determined until they mate). Traits evolve in individual animals, because they help control conditions that affect the reproduction of the mating pair. Mating itself is the most obvious example of behavior that serves this function, but it includes, indirectly, at least, all the “sexually selected” traits required to mate, from suitable organs for the mating process to brilliant plumage in some male birds and fights over mates among male herd animals. Such genes can evolve, like any gene, because of their contribution to the maximum holistic power of the reproducing organisms of which they are part. It is just that the reproducing organisms of which they are part are the peculiar, disjointed primary structures of the kind entailed by sexual reproduction.
After the evolution of sex, therefore, what is evolving might be called a “composite organism,” or "composite primary structure," for the unit of selection is composed of more than one individual organism. And after the evolution of multicellular animals, the individual organisms may be in different generations. Reproduction is carried out by specialized cells, and thus, the mating pair does not necessarily die in reproducing sexually. Multicellular animals may, therefore, live side by side with their offspring, and when reproduction comes to depend on offspring being nurtured and protected by parents, the composite reproducing organism includes the whole family. The family is the primary structure that goes through the reproductive cycle (which is the ontological cause of gradual evolution), and traits evolve in its members because they control conditions that affect the reproduction of the primary structure as a whole.
Altruistic genes can evolve, like other genes, because they are structural causes of behavior in one, disjointed part of the primary structure, and they control a condition that affects the reproduction of the whole primary structure by promoting the reproduction of another individual organism of which the whole is composed. The disposition of children (and even grandchildren) to sacrifice for one another is just another way, along with caring for offspring, in which the same disjoint primary structure does the non-reproductive work that controls all the conditions affecting its reproduction as a whole. Like any gene, altruistic genes are working together with all the other genes of an organism to control all the conditions that affect its own reproduction as a whole. It is just that the primary structure, or reproducing organism in this case, is a collection of distinct multicellular animals.
Reproductive causation offers, therefore a ready explanation of altruism in the family (as an ontological explanation, if you will, of so-called “kin selection” on the “selfish gene” theory). But that is not quite an adequate explanation of the kind of altruism required for the existence of nomadic bands of hominids, because their protective altruism must extend beyond members of the family to primates that are not kin. Several families must band together in order to have enough adults for the protection of everyone, and in the long run, inbreeding among them is ruled out by its deleterious effects. As sociobiologists have noted, primates had already evolved mechanisms to prevent inbreeding, such as the disposition of young males to leave their home group at sexual maturity and seek mates in other groups, and hominids without some mechanism for spreading their genes beyond the nomadic band would not have been successful for long.
If strangers, rather than kin, are benefited by animals who sacrifice themselves altruistically, altruistic genes will make strangers who lack the altruistic gene more likely to reproduce, and that will make the primary structure (or “reproducing organism”) of which the altruistic individuals are part less likely to reproduce, given the scarcity of resources caused by population growth. A family of altruistic animals is likely to be invaded by “free riders,” who benefit from altruism without reciprocating, and altruism will be extinguished.
It is possible, of course, to conceive of mechanisms that would extend altruism beyond the family. For example, if altruism happened to be accompanied by a “tit-for-tat” policy of aiding only others who reciprocate, but not aiding those who do not, a group of altruistic animals would be able to resist invasion by “free riders.” Computer simulations suggest that such conditional altruistic behavior would enable altruism to evolve beyond the family. But to explain the evolution of an altruistic trait in a whole species in this way would require a series of different selection pressures whose origins remain unexplained. The altruistic gene must first evolve by kin selection in groups made up only of family members so that the “tit-for-tat” condition can be added later when for some reason family groups are joined by strangers. This is just the sketch of a possible explanation which depends on unexplained changes in the environment. In order to explain altruism among hominids, it would be necessary to fill in the concrete details of human evolution. And in the end, it would be merely a “just-so” story, which makes the evolution of hominids an accident.
There is, however, another explanation, which entails that the evolution of hominid altruism is inevitable. It is not an accident that the radiation of manipulative animals includes some that invade the grassland, because manipulation is only likely to evolve in animals living in trees. And their new ecological niche imposes a form of group selection on the nomadic primates. In other words, there is a primary structure, or "composite organism," that is even more inclusive than the family and is evolving by reproductive causation (though in this case, there are actually two units of selection (or primary structures), because the individuals or families continue to evolve by reproductive causation within them).
Under the conditions in which nomadic primates found themselves, there would be at least a weak form of group selection, and it would inevitably become stronger. What is needed is a disposition to protect other members of one’s nomadic band from predators even when the others are not members of one’s own family. Groups having some members who happened to be disposed to fight off predators, regardless who in the group was attacked, would be the groups that were most likely to succeed in acquiring energy-rich objects from the grasslands. Thus, altruistic protection of others from predators would evolve in individual primates.
The selection pressure at the group level was probably not very strong at first, because nomadic groups would tend to disband and regroup rather fluidly when they met or returned to forested regions where they were safe from predators. Groups that happened to have families with courageous altruists would be more likely to return whole, and that would tend to favor the altruists. The desire to carry clubs and protect others in one’s group would tend to evolve.
Group level natural selection would, however, become more powerful, because they would be less likely to disband. Groups with altruistic protectors would be more powerful in controlling relevant conditions as they formed bonds that made them stay together, because they would have established habits and routines that worked together more effectively in protecting against predators and controlling other conditions. And as groups became more powerful and more permanent, group level selection would become stronger, for they would eventually impose natural selection on themselves at the group level. Not only would the populations of those bands increase faster than those of other primates venturing into the grasslands, but the groups themselves would reproduce. Groups must divide up as their populations grow, because there is a limit to how many members can be sustained by the energy acquired from the amount of land that can be covered by wandering around. Hence, there would be organisms (or primary structures) going through reproductive cycles at two levels of organization at once: at the individual (or family) level and at the level of nomadic bands. And the reproducing organisms (or primary structures) on both levels would impose natural selection on themselves by their population growth and the scarcity it causes. Thus, while individuals continued to evolve functional traits that made them increasingly powerful as members of such groups, the groups themselves would evolve traits that made them increasingly powerful in controlling conditions which affect the reproduction of the group as a whole. That would include many traits in addition to the desire to protect other members of one’s group.
 Limits
of Social Coordination. Although motivating social, or even altruistic,
behavior is not, therefore, an obstacle to the evolution of nomadic bands
of hominids, there is nevertheless a severe limit on their power, because
they are still just animal societies. This can be seen by analogy to multicellular
animals.
Limits
of Social Coordination. Although motivating social, or even altruistic,
behavior is not, therefore, an obstacle to the evolution of nomadic bands
of hominids, there is nevertheless a severe limit on their power, because
they are still just animal societies. This can be seen by analogy to multicellular
animals.
On the individual level, animal power comes from two sources. One source is the size, shape, and strength of the body, and the other is the structure of its animal behavior guidance system. Both depend on its biological behavior guidance system, although the power of animal behavior comes from the geometrical structure it imposes on the motion and interaction of material objects in the region. We have seen how the animal behavior guidance system enables the animal to act on objects in space so that it structures the thermodynamic flow of matter from potential energy to evenly distributed heat.
On the social level, the power of a group has comparable sources. The size of its population corresponds to the strength of the body, and the mechanism for coordinating behavior corresponds to its behavior guidance system. Although in some ways, it is easy for the group to act as a whole on an object in space, say, by chasing it, there is a certain kind of structure that animal societies are not able to impose on the thermodynamic flow of free energy toward increasing entropy, even though it would make them more powerful. It is the coordination of members' behavior that would generate social level behavior like the behavior generated by manipulative animals. That is the function that explains the evolution of language.
What makes behavior animal behavior is that it acts on other objects in space, either to ingest energy rich object or for other purposes, and the function of the animal behavior guidance system is to adapt the spatial aspects of behavior to spatial aspects of the situation so that it controls the relevant conditions. Since the animal itself uses free energy to do work, the generation of bodily behavior is already one way of imposing a geometrical structure on the thermodynamic flow of matter in the region, and so when the body moves around in space and interacts with objects throughout the region, it imposes a further geometrical structure on the thermodynamic flow in the region. Manipulative animals are able to structure the thermodynamic flow even further by coordinating the behavior of its hands, and that points to the potential power of animal behavior at the social level: manipulative animal behavior is possible at the social level if the animal behavior of the members of the group can be coordinated.
Animal societies. By “animal societies,” I mean societies of animals, from schools of fish and flocks of birds to herds of ungulates and packs of wolves. Animal societies are so weak at the social level that they are not really animals at all. Groups of animals do have some power to control conditions in the world. It comes from the power of its members, and it can be increased up to a point by adding new members to the animal society. But animal power depends, because of the nature of animals, on adapting the spatial aspects of social level behavior to the spatial aspects of the situation, and that does not happen naturally in groups of animals. It requires a structural cause of some kind, that is, a social level animal behavior guidance system. Thus, the question is, Where is the material structure that would enable an animal society to behave like an animal on the social level? Or like a social-level manipulative animal.
The tendency toward randomness applies to animals like other objects that move and interact in the region. Solitary animals stay out of one another’s way, for the most part, and forage on their own, or, at most, in family units, so that the way their behavior adds up in space over time tends to even out their distribution in space.
To say that they are solitary is not to say that they are not social at all. Solitary animals often stake out territories from which to gather the energy they will need, and they defend it against conspecific intruders by a more or less ritualized fighting behavior.
Animal societies do exist, however, because some animals have instincts or desires that act like a force attracting animals of the same species to one another. Gregarious instincts are like gravitation, making the motion and interaction of such material objects adds up in space over time to the formation of social level objects in space.
But gregariousness results only in the minimal geometrical structure required to be a composite object in space, resembling gravitational objects, such as stars or drops of water, more than the kinds of material objects that can serve as structural causes. The disposition of fish to swim in schools and ungulates to run in herds are examples of such “gravitational” organizations at the social level. Though the members do not get very far apart, the spatial relations among the animals are continually changing at random.
In order to be a social level animal, the members of an animal society would have to act on objects at the social level like animals do at the multicellular level, and more power would come from acting like manipulative animals at the social level. Gregariousness itself is, perhaps, a way of acting as a whole. But its social level behavior is hardly up to the standard set by multicellular animals
In herd animals, the members’ desires to stay relatively close to one another includes the desire to start running when other members do. Thus, if the members also have the desire to run away from any predators they may spy, the whole herd will have protection from predators. Herd animals can usually run long and fast, and so as long as they run together, some members of the herd will avoid predation by outrunning any predators they may encounter. This is presumably the function of the herd instinct.
Leadership can make merely “gravitational” social level behavior more functional. Herd animals do not need a leader, because that role can be played by any member that happens to spy the predator first. But leadership can be useful to carnivores in acquiring energy. Wolves, for example, can all reliably attack the same animal, if they have a leader. Leaders are provided by dominance hierarchies. With the right kinds of conditional, instinctive desires, a competition or face-down among members can determine a “pecking order” among the members. The selection of goals for social level animal behavior is then centralized in a leader, and they can prey on animals that would be out of reach for individual animals, not to mention making it possible to settle conflicts among members over food or mates.[1]
Locomotion is about the limit of social level behavior in animal societies, because the other aspects of animal behavior, such as attacking prey and eating, are left up to the individuals acting in parallel. Societies of subjective animals are inherently more powerful than societies of telesensory animals, since they are less dependent on the inheritance of rigid instinctive routines for individual animal behavior. And group level locomotion can be more powerful with a higher level of neurological organization to guide it. Packs of wolves make better use of group level locomotion than flocks of birds, and groups of dolphins can use locomotion for purposes that are beyond schools of fish. But even societies of manipulative animals can only act in parallel with one another. Elephants, for example, may join forces in tearing down a tree, but even if they are manipulative animals, they cannot intentionally fold a tree into some shape like two hands working in different ways to produce a single result.
Need for social coordination. To put it positively, in order for animal societies to have animal behavior at the social level that is analogous to the behavior of even multicellular animals, it must be possible for the members to divide up tasks that work together as whole like different parts of an animal body, such as jaws working against one another to catch prey. This would be especially valuable for groups of hominids in acquiring food, since they cannot run fast enough to prey on herd animals like predatory carnivores. For example, if the group could make some members chase herd animals down a ravine where others were waiting to club them or roll rocks onto them, the group could kill animals that would be out of reach simply by chasing them. It would enable hominids to tap sources of energy that previously were available only by scavenging leftovers from carnivores.
The key to generating animal behavior at the social level is, therefore, coordinating the behavior of the members in a sense that involves a “division of labor” among the members. That would make the animal society more like the body of a subjective animals, for the animal society as a whole could adapt the spatial aspects of its social level behavior to the spatial aspects of the situation, much as different parts of a multicellular body are coordinated by motor commands to act as a whole on other objects in space.
Furthermore, such coordination of the members’ behavior in group action would be even more powerful, if the “motor commands” to different parts of the body were not merely instinctive, nor even discovered by trial and error, but designed in structural imagination to adapt the spatial aspects of social level behavior to the spatial aspects of the current situation, like primates coordinating the movements in different hands to attain a single goal.
Social coordination by the biological behavior guidance system. It is possible for the division of labor among the members on which coordination depends to come from the biological behavior guidance system, because the goals that animals pursue depend on the desires that it builds into them. But the biological behavior guidance system is not much help, because it cannot divide up labor very much, except by crippling social level animal behavior in another way.
In animal societies. It is possible for the biological behavior guidance system to give different members of the same species different desires, because in sexually reproducing multicellular animals, as we have seen, the unit of selection can be the entire family. When the organism going through cycles of reproduction is not just the individual, but the mating pair and its offspring, their behavior can be naturally selected for controlling conditions that affect the reproduction of the whole organism. Males and females can be given different desires.
In some African wolves, for example, the females have the desire to stay in the den and care for their young while the males have desires that lead them to hunt together in a pack, eat their prey, and return to the den and regurgitate food for their families.
More common are mating rituals and elaborate, ritualized fights by which males determine which of them will have access to females.
Likewise, the biological behavior guidance system can give parents and children different desires. We have seen how the peculiar disjointed nature of these “composite organisms” (or composite primary structures) can account for altruism among children and grandchildren.
Age and sex differences do not, however, afford enough division of labor to enable animal societies to act as social level animals. And even if it did, it would not divide up labor differently in different situations, because instincts are shaped only by natural selection of whole biological behavior guidance systems.
The maximum coordination in animal societies provided by the biological behavior guidance system is illustrated, perhaps, by the wild dogs of Africa. It is said that they can herd other animals in certain directions, using barking to coordinate their behavior, so that other dogs waiting for them can ambush the prey as they run by. This improbable geometrical structure in their social level behavior can trap their prey, like the limbs of a single animal body reaching out to pull energy-rich objects into its jaws, even though it is just how the motion and interaction of independent bodies add up in space over time because of the desires that motivate them. This is to add group level ingesting of energy rich object to locomotion in the repertoire of animal societies.
But like all instinctive behavior, it depends on the biological behavior guidance system, and thus, it still cannot be adapted to new situations except by the relatively slow process of biological evolution.
In multisomatic animals. There is one way that the biological behavior guidance system can impose a more complex division of labor. It occurs, as we have seen, in multisomatic organisms, such as insect colonies, where in addition to age and sex differences, members are determined to have different kinds of behavior by the pheromones, or messenger molecules, they encounter.[2] But this mechanism of coordination requires animals to have specific locations in the geometrical structure of the whole organism in order for them to encounter the right pheromones at the right times. The unchanging geometrical structure is provided by a hive or labyrinth of tunnels. The pheromones are distributed to specific locations in the process of setting up the multisomatic organism, and in at least one mammal society as well as many insect colonies, it centers on a queen whose offspring populate the colony.
This possibility is exemplified by the blind mole rat of East Africa. Their reproduction is organized around queen, which is located at the center of a network of tunnels in the ground, and pheromones are distributed among them, determining different members to pursue different goals, by how they eat one another’s feces.
There is, in other words, a structural cause of animal behavior on the social level in multisomatic animals. But it is of no use to animal societies, because they have no unchanging spatial relations among the members. They are merely “gravitational objects” on the social level, sustained by gregarious desires, with the spatial relations among the members continually changing. To acquire a geometrical structure as a whole would constrain the motion and interaction of its members severely, for they would be sedentary and able to act only like moving parts in an otherwise rigid machine.[3]
As mere animal societies, therefore, bands of hominids had only a limited capacity to generate social level behavior. The radiation of these primates into the grasslands made them bipedal. They carried clubs and may even have wielded axes to fend off predators that attacked them, including members other families. These traits were inevitable, and adaptation to membership in nomadic bands may account for other hominid traits. Though dominance hierarchies resolved conflicts and provided leadership, the need to remain in proximity to one another strengthened and refined social desires. And being manipulative animals, they could accumulate and pass on useful skills and habits (especially if they had the capacity to imitate one another’s behavior). As skill at wielding clubs increased, other tools were probably added, making it possible to hunt and kill larger game. They may have evolved group level hunting behavior as complex as wild dogs, and their structural imagination may have given them more complex signals for coordinating their behavior. They may even have learned to control fire. And the group level natural selection that nomadic bands imposed on themselves by the scarcity caused by their own population growth may have favored neurological refinements that account for the expansion of the brain. But since those nomadic bands were still just animal societies, hominids encountered the same limit on their power to generate social level behavior as other animal societies.
 The
original function of natural sentences. Though the evolution of wild
dogs and societies made up of other merely subjective animals had to stop,
once reproductive causation had made them as powerful as possible, the evolution
of hominid society would begin a new stage of evolution, because primate imagination,
with its capacity to understand structural causation, made a revolutionary
change possible. That change was a new kind of animal behavior guidance system
at the social level. But it was unlike the structural causes that had evolved
to guide behavior at lower levels of biological organization. It was a primitive
language.
The
original function of natural sentences. Though the evolution of wild
dogs and societies made up of other merely subjective animals had to stop,
once reproductive causation had made them as powerful as possible, the evolution
of hominid society would begin a new stage of evolution, because primate imagination,
with its capacity to understand structural causation, made a revolutionary
change possible. That change was a new kind of animal behavior guidance system
at the social level. But it was unlike the structural causes that had evolved
to guide behavior at lower levels of biological organization. It was a primitive
language.
Though in animal societies, social level behavior is never anything more than how the animal behavior of members with different desires (and, thus, different intentions or goals) add up in space over time, language is a mechanism that reaches inside each individual’s behavior guiding system and controls its animal system of representation. By connecting all these innermost mechanisms, language can give all the members of the group a single, common intention, and thus, even though they may all be performing different tasks, the group acts on other objects in space as a whole.
The reason that hominid society could evolve into spiritual animals, while other animal societies, such as packs of wolves and herd of ungulates could not, was structural imagination. Language requires a higher level of neurological organization, and it could be tried out as a radical random variation only on hominid brains.
How natural sentences coordinate behavior. Only a primitive language is needed to guide social level behavior. By a “primitive language,” I mean a language with “natural sentences,” that is, sentences with a simple subject-predicate grammar (or subject-verb-objects grammar). Natural sentences can refer to individual members and represent them as behaving in certain ways (or in certain ways in relation to certain other objects in space), and thus, a set of natural sentences, one for each member, can represent a plan for social level behavior by assigning different roles to different members. Here is how it would work.
In the beginning, the leader of the group would imagine a way of coordinating behavior in the situation that seemed to satisfy their desires, and he would assign roles to individuals by using natural sentences, in everyone’ presence, to describe how each member was supposed to behave. That public linguistic interaction would give the members a shared understanding of what every member was supposed to do in advance of acting. That is, each member would know not only what he was supposed to do, but what everyone else was supposed to do. Since members could play complementary roles in a common plan, the society could act as a whole on other objects in space and thereby control relevant conditions that were previously out of reach.
Coordinating behavior was not necessarily as simple as a leader telling each member what to do, because the use of natural sentences depends on imagination, which is a form of understanding by which animals can see what is possible. The manipulative animal system of representation uses images and how they change over time to represent regularities about change that are caused by the motion of objects relative to others and by the manipulation of objects, and since public utterances can control everyone’s imagination, every individual would be able to “see” the difference that each member’s behavior would make. By fitting those individual contributions together in “mammalian space,” each member could see what consequences they would have jointly in the situation thereby understanding how their behavior would attain some goal. The coherence of the plan would be tested in the process of adopting it, because each individual would understand what his own behavior was supposed to contribute to the plan and they would not be as inclined to follow the leader, if the whole plan or their role in it did not make sense to them.
Analogy to the multicellular biological behavior guidance system. There is a useful analogy to the multicellular biological behavior guidance system. The use of natural sentences to coordinate the behavior of individual animals resembles how the multicellular biological behavior guidance system coordinates the behavior of cells in constructing the bodies of multicellular (and multisomatic) animals. In both cases, a copy of the entire plan for the higher level structure is distributed to each lower level organism. Each cell receives a copy of the entire genome by the asexual reproduction of the fertilized egg cell in the process of embryological development, and each member of the spiritual animal receives the same plan of coordinated behavior by public, linguistic interactions in which the leader tells each member what to do. And in neither case will this means of coordination work, unless each member receiving a copy of the plan identifies itself with a different role in the plan, for otherwise it will not be able to generate the special kind of behavior required of it.
In the multicellular biological behavior guidance system, each cell can identify with a certain part of the genetic blueprint for the multicellular body because each cell has a fixed location relative to all the others in the process of embryological development. Different cells are determined to open up different segments of their chromosomes and generate different kinds of behavior by their locations in the geometrical structure of the body as a whole and the messenger molecules received there. (In insect societies, the process of development typically uses a “queen” to produce most of the multicellular members and construct a hive, and different members are determined to behave in different ways by the pheromones located where they find themselves in that structure.)
With the use of natural sentences, an unchanging geometrical structure is not necessary, because the plan is distributed by public linguistic interactions. The members all have animal systems of representation by which they are aware of the locations of one another’s bodies and other objects in the region, and thus, when the leader assigns different roles to different members, not only do the members construct the same plan of behavior relative to the same objects in space in their own faculties of imagination, but the members also identify each member with a certain agent in the plan, including themselves. Thus, different members intend to behave differently when the time comes, but their actions all fit together as a whole.
Contrast to animal signals. Even a primitive language is fundamentally different from the communication used in animal societies. Cries and screams of various sorts are used by other social animals, but such signs do not control imagination in the same way as natural sentences. Signals do enable them to communicate with one another and affect one another’s behavior, but they are inherently incapable of assigning tasks to members of a group.
Wild dogs of Africa use a system of different kinds of barks to coordinate their behavior in herding other animals toward the kill, and honey bees perform a dance by which those returning from a source of energy can indicate its direction and distance to other bees that are just departing. Such animal signals can be versatile, communicating different messages in different situations. As many as sixty different cries have been identified in societies of monkeys, and the hominid mouth and larynx were probably shaped by reproductive causation to generate many more kinds of signals.
But signaling cannot be used to assign tasks to individuals in advance of generating social level behavior, because signals and signs depend for their meaning on the locations of the animals sending and receiving them. Honey bees can communicate the distance and direction to the source of energy, but only because the other bees emerging from the hive already have the goal of traveling to a source of energy and they know to take the directions for locomotion as relative to the hive, the stationary body of the multisomatic organism where the signals are given. Monkey cries are also relative to the location of the signaler, for example, indicating where a certain kind or predator has been detected or where help is needed. Likewise, barking may coordinate the behavior of wild dogs relative to one another so that together they herd other animals in a certain direction, but the goals are fixed and the messages are part of an instinctive pattern that has been shaped to have this effect by reproductive causation.
This dependence of animal signals on the context of their utterance for the communication of their references and meanings makes them useless in assigning different roles to individual members in a plan of social level behavior. In order for animals to use verbal behavior to share a plan that divides up tasks among them, it must be possible, in advance of acting and regardless of its context, to refer to particular members and describe how each is to behave at a later time so that every member can understand who is to behave how and in what situations. That is what the simple grammar of natural sentences makes possible, and since animal communication lacks a grammatical structure, other animals do not have even a primitive language.
Contrast to animal societies. Language makes spiritual animals fundamentally different from animal societies, because the use of natural sentences is a structural cause for social level behavior. Animal societies have no structural cause for their social level behavior. Their social level behavior is simply how the behavior of members with possibly different desires (and, thus, different intentions or goals) add up randomly in space over time. The members act alongside one another, each pursuing its own goal.
The use of natural sentences is, however, a structural cause of social level behavior. Though there are no unchanging spatial relations among the members, they are in more or less continual linguistic interaction, and their linguistic interactions distribute to every member a plan for social level behavior toward some object in space in which each member is assigned a specific task that is designed to fit together with other members so that the group as a whole attain some goal. The result is a form of animal behavior, because it acts on other objects in space. That is, the group as a whole channels the thermodynamic flow of matter toward evenly heat to do work by imposing a geometrical structure on the region in ways that control conditions that affect their reproduction. Their social level behavior is a structural global regularity, and it has a structural cause, even though it is not a simple material structure.
The structural cause is the use of natural sentences, because that is what distributes the same plan of joint action to all the members, including the special instruction that informs each member about his special task. Since the behavior of all the members is represented in the imagination of each member prior to acting, the members of spiritual animals can be said to have the same intention. Instead of pursuing private goals alongside one another, they are all pursuing a single, shared goal. That is the sense in which the spiritual animal itself may be said to intend to behave in the way described by the plan.
That is, the spiritual animal works like a machine. With each member of the group playing a different role in their common plan, each member is like a moving part of an invisible machine whose motion and interaction is constrained by its location in that invisible geometrical structure of the whole machine. But what makes the machine invisible is that its structure as a whole is just the shared plan. The plan requires only that each member be able to imagine how all the participants will move and interact in the situation as a result of exchanging linguistic representations and act accordingly, and thus, the machine's structure is not only invisible, but infinitely flexible.
But since groups of language using primates have no body apart from all the independently moving bodies of its members, the social level animal can be said to have spiritual body. That is the reason for calling it a spiritual animal.
Contrast to multisomatic animals. Multisomatic animals, such as insect colonies, also have social level behavior generated by a structural cause, but it depends on having a physical body at the social level and that makes them less powerful than spiritual animals.
Since the unchanging geometrical structure of the multisomatic body comes from a hive or network of tunnels, their physical body makes them sedentary. However, the spiritual animal is not sedentary. It can move around as a whole and act on other objects in space like a manipulative multicellular animal on the social level of biological organization. It can, in effect, send different motor commands to different parts of its body, except that, with a spiritual body, there is no constraint on how each part can move and interact in relation to one another. It is as if its hands could manipulate objects without arms connecting them to the body.
Furthermore, in multisomatic animals, the coordination of members’ behavior is like an inherited instinctive routine, because it is determined by their biological behavior guidance system. The spiritual animal, on the other hand, has the use of the most highly evolved animal system of representation at the multicellular level to coordinate its behavior. It does not have to wait for instincts to evolve in order to adapt spatial aspects of its behavior to spatial aspects of the situation, as telesensory animals do. As long as the members have a desire to do what the leader instructs them to do, it does not even have to rely on trial and error to shape its social level behavior to fit situations, as subjective animals do, because structural imagination can be used in advance of acting overtly to design social level behavior so that it will control relevant conditions by how it fits together spatially with the other objects in the current situation. Different “motor commands” are sent to different parts of its spiritual body in the linguistic interaction by which the leader explains the plan. Thus, spiritual animals can generate novel, but appropriate kinds of animal behavior in the situation — much as Kohler’s ape did in using a stool and stick to knock down the banana, except on the social level.
![]()