![]()
 Animals.
Though the evolution of multicellular plants may also involve a series of
stages, nothing more will be said about them here, because the current project
is to show that there is a series of inevitable evolutionary stages leading
up to rational beings like us (and beyond). The evolution of multicellular
plants is a separate branch of evolution. But to lay the foundation for tracing
the evolution of animals through a series of inevitable stages to rational
beings like us, let us first consider the basic nature of animals and inventory
the kinds of animals that can evolve, both basic and anomalous.
Animals.
Though the evolution of multicellular plants may also involve a series of
stages, nothing more will be said about them here, because the current project
is to show that there is a series of inevitable evolutionary stages leading
up to rational beings like us (and beyond). The evolution of multicellular
plants is a separate branch of evolution. But to lay the foundation for tracing
the evolution of animals through a series of inevitable stages to rational
beings like us, let us first consider the basic nature of animals and inventory
the kinds of animals that can evolve, both basic and anomalous.
That will leave us to consider, in subsequent chapters, the differences among animals that depend on in levels of neurological organization at the multicellular stage. The social level of biological organization will be introduced by way of insect colonies, for they are an example of the anomalous animal that will be explained in this catalogue. Though human society is also an animal on the social level, it depends on the stages of animal evolution caused by levels of neurological organization, and thus, there is not much to say about it until after we trace the stages of animal evolution.
 The
Nature of Animals. The difference between animals and plants (and plant-like
organisms) is part of the differentiation of organisms into kinds that occurs
at each stage of biological evolution. What makes animals different from plants
is not their level of part-whole complexity, but that animals acquire the
free energy (and parts) they need to fuel their reproductive cycles by ingesting
other objects in space. Those objects are not mere photons, but material objects,
with rest mass — sometimes just energy-rich molecules, but usually other organisms.
The
Nature of Animals. The difference between animals and plants (and plant-like
organisms) is part of the differentiation of organisms into kinds that occurs
at each stage of biological evolution. What makes animals different from plants
is not their level of part-whole complexity, but that animals acquire the
free energy (and parts) they need to fuel their reproductive cycles by ingesting
other objects in space. Those objects are not mere photons, but material objects,
with rest mass — sometimes just energy-rich molecules, but usually other organisms.
Animals require, therefore, a kind of behavior that is radically different from anything plants require. In order to ingest other objects in space, animals must have a kind of behavior that acts on them, whereas plants (and plant-like organisms) need only behavior that acts on the world as a whole at the moment. Though plants often need to orient their bodies and their behavior in the world, they act, at most, only in some direction relative to a field in space, such as gravitation, in the direction of electromagnetic radiation, or relative to gradients of water in the soil, the magnetic field, and the flow of air or water. Such behavior is not directed toward particular objects in space, but merely affects whatever objects may be located in the direction in which its behavior is oriented. By contrast, no animal, regardless how simple, could not control even the most basic condition that affects its reproduction as an animal, unless it could direct its behavior toward particular objects in space. Thus, we might say that, whereas plant behavior acts on the whole world at the moment (or, at most, some direction in space), animal behavior acts on objects in space.
A new system is required to guide the animal’s special kind of behavior, because ingesting other objects is a hazardous way of making a living. Ingestion is an intimate way of relating to another objects, and since there are usually many kinds of objects around that would harm the animal, if ingested, animals must have a mechanism of some kind by which to tell in advance which objects to ingest and which to pass up. Eating and not eating are incompatible behaviors with respect to any given object in space, and selecting which objects to eat is, therefore, a condition of the existence of reproductive cycles fueled in this way. Such a basic function may well be called a "need," and thus, needs, like functions, can be used to explain the traits that evolve.
That is, there are various ways of doing non-reproductive work within the range of random variations being tied out, and those that make it possible for organisms to acquire energy as animals do will be selected by their power to control that relevant condition. Thus, reproductive causation implies that organisms with animal behavior guidance systems will evolve, if they are possible.
The animal behavior guidance system is a second system for guiding behavior in most animals, because as living organisms, animals already have a system that selects between incompatible kinds of behavior, namely, the biological behavior guidance system. Such a mechanism is needed to coordinate the behavior of the lower level organisms that are bundled together as the higher level organism, and it is responsible for guiding the organism’ through its reproductive cycle.
The first such biological behavior guidance system was at the prokaryotic level. It was necessary, as we have seen, because the two kinds of work that the first organisms had to perform during each cycle in order to evolve, growth and reproduction, were incompatible and could not be generated at the same time. Its origin during the prokaryotic stage marked the beginning of life, since the ability to select and generate both kinds of behavior is what enabled organisms to go through reproductive cycles on their own, independently of the cycle of night and day. That gave them the autonomous activity ordinarily meant by saying that something is alive, and as living organisms, that make themselves evolve.
At each subsequent stage of biological organization, a new biological behavior guidance system was needed, because it was necessary to coordinate the behavior of the new kind of lower level organisms that are bundled together and go through reproductive cycles as a whole. Since, at any biological level, animals already have a biological behavior guidance system, and he animal behavior guidance system is their second system for guiding behavior, one whose function is to guide the special kind of behavior that animals need in order to acquire free energy by ingesting other material objects in space.
The reason for calling both mechanisms "behavior guidance systems" is that they similar kinds of functions. In both cases, a basic choice must be made between incompatible kinds of behavior in order for organism of their kind to go through reproductive cycles at all. For organisms to be alive, it is the choice between growing and reproducing, whereas for living organisms to be animals, it is the choice between eating a particular object and not. There is no way to avoid making such a choice in either case, for the two essential kinds of behavior cannot be generated at the same time. (It is less inconvenient in multicellular organisms, with their unique way of reproducing, because this choice is made when cells are determined to have different functions and some are assigned the task of reproducing the organism as a whole.) Since the biological and animal behavior guidance systems evolve to serve similar functions, we can discern a basic similarity in their structures, because they must serve similar subfunctions.
A behavior guidance system is not a mere cybernetic device, as we have seen. Cybernetic systems use feedback from the consequence of their behavior to attain or maintain some overarching goal. But behavior guidance systems are essentially different, because their function is to select between incompatible goals. The difference can be seen in how the behavior guiding function requires mechanisms to serve no less than three sub-functions, regarding input, selection and output.
Input. In order to make the selection among possible goals, input is required about the current state of affairs, either about the world as a whole or about particular objects in space. That means that the system must be affected by the state of the world (including, perhaps, the state of the organism itself) in a special way when choosing one of the alternatives or the other would control the condition that affects reproduction.
Selection. The input must change a state of the behavior guidance system in some way so that the system selects the right goal from the alternatives, which is its basic function. This is the sub-function that a cybernetic system lacks.
Output. In order for its selection to make a difference, the behavior guidance system must also be able to generate the appropriate kind of behavior. Since it is directed at a goal, such output typically has the structure of a cybernetic system, using feedback from the effects of earlier behavior to adjust its output to attain the chosen goal.
Thus, the behavior guidance system's three essential sub-functions are receiving input, selecting the goal, and generating output (which may also use input as feedback). It cannot get by with fewer sub-functions, whereas the cybernetic system, with a fixed goal, requires only a mechanism for registering input and generating behavior accordingly.
The nature of reproductive causation entails, as we have seen, that non-reproductive work is not a means to reproduction as an end in the biological behavior guidance system. The goals of non-reproductive work are the conditions affecting reproduction that must be controlled, whereas reproduction is merely the cause of the natural selection by which they evolve.
Animal behavior itself is different from the kinds of behavior guided by the biological behavior guidance system, because it must be directed at other objects in space, not merely at the world as a whole (including orientation in a field of some kind). The spatial aspect of animal behavior may not be obvious in some cases. It can be just an aspect of the unchanging structure of the animal body as, for example, in sedentary animals that filter the flow of water to feed on smaller organisms. But the spatial aspect is essential, because the free energy that animals use is contained in objects that have locations in space. Those objects must be ingested, and in order for animals to become more powerful, they must acquire traits that take advantage of how their sources of free energy are located in space.
Though the behavior guided by biological system is not directed at any other particular objects in space, but only at the world as a whole, it can still gradually evolve greater power, because it can organize different kinds of behavior in time. As we have seen, the biological behavior guidance system is a higher level structural cause that coordinates the behavior of the many lower level organisms of which it is composed, and it is responsible for all the behavior required to complete a whole reproductive cycle, including both growth and reproduction. Such a complex material structure is what made it possible to explain gradual evolution as a global regularity that is caused ontological by reproductive cycles and the wholeness of space.
Though the cycle of reproduction does not exist fully at each moment, its structural cause does, and since the entire cycle is implicit in a material structure (that is, its biological behavior guidance system) which does exist fully at each moment, the ontological cause of gradual evolution could be described, paradoxically, as a four dimensional object that endures through time and reproduces in space. Thus, implicitly contained in the structure of the biological behavior guidance system are not only both the phases of growth and reproduction, but all the responses to changing circumstances and sequences of behaviors using feedback to attain goals that occur during the entire reproductive cycle.
It may be surprising that nothing but the right sequence of activities at the right times is needed to generate a whole reproductive cycle (except, perhaps, for orienting itself in various fields). But taking advantage of the power to sequence behavior temporally is what enables the biological behavior guidance system to become increasingly powerful at each stage of evolution. In addition to making each behavior as effective as possible in controlling its relevant condition, reproductive causation inserts or deletes kinds of behavior from temporal sequences, changes their duration or quantities, and rearranges their sequences in time. The right sequence (conditioned on relevant input) can be very powerful. That is why the biological behavior guidance system is the locus of most evolutionary changes in such organisms.
This difference between animal and merely biological behavior is ontologically basic, because in addition to any temporal sequencing of behavior that is generated by the biological behavior guidance system, the animal behavior guidance system also imposes a spatial structure on the behavior of the organism by acting on other objects in space. Insofar as the power of the animal behavior guidance system to control what happens increases by taking advantage of the fact that the objects containing free energy have locations in space, animal behavior is superimposing a geometrical structure on the region.
Organisms can evolve only in regions where there is a supply of free energy, and as we have seen, there is a region-wise geometrical structure about the thermodynamic flow of matter from potential energy through kinetic energy (and radiation) toward evenly distributed heat that constitute free (or usable) energy. Thus, we can see animal behavior as channeling the thermodynamics flow in the region. In other words, what most material structures do simply by existing in the region, animals do by acting on objects in space. That is what makes animals more powerful than plants, and it suggests how far animals might evolve in the power to control relevant conditions.
Structural causes generate irreversible structural global regularities by using free energy to do work, that is, making things happen that would not otherwise happen. Though animals are just organisms contained along with other objects in the region, the evolution of behavior that takes into account the locations of other objects in space in order to control conditions that affect their behavior can channel the flow of free energy toward increasing entropy over very large regions of space. In other words, the spatial aspect of animal behavior (being directed at objects in space) imposes a geometrical structure on the region (and, thereby, channels the thermodynamic flow in the region in a way that would otherwise require a material structure as large as the relevant region, like a giant machine or building). The actualization of the potential of animal behavior to do work in the region is what we are tracing by deriving from spatiomaterialism the evolutionary stages that determine the overall course of evolution.
This potential is actualized mainly in multicellular animals because of the evolution of a series of level of neurological organization in them, but before we trace those stages, let us catalogues all the other animals to see how multicellular animals are related to all the other kinds of animals.
 Kinds
of animals. Stepping back from the multicellular level of biological
organization, this explanation of the nature of animals entails a classification
of animals into their natural kinds. The basic kinds of animals are easy to
classify, because they are ontologically necessary, and identifying them will
provide a map of subsequent stages. However, this way of sorting out the basic
kinds of animals resolves several puzzles about the classification of animals
in biology, and thus, we will conclude with a survey of the kinds of anomalous
animals that are just as ontologically necessary.
Kinds
of animals. Stepping back from the multicellular level of biological
organization, this explanation of the nature of animals entails a classification
of animals into their natural kinds. The basic kinds of animals are easy to
classify, because they are ontologically necessary, and identifying them will
provide a map of subsequent stages. However, this way of sorting out the basic
kinds of animals resolves several puzzles about the classification of animals
in biology, and thus, we will conclude with a survey of the kinds of anomalous
animals that are just as ontologically necessary.
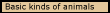 Basic
kinds of animals. Since animal behavior guidance systems evolve at
each level, the most basic divisions among animals correspond to the levels
of part-whole complexity in the biological behavior guidance system discussed
in the last section. We should expect some kinds of animals at each level
of biological organization, because animals are just a variety of the living
objects that evolved at each stage of evolution. They are, in technical terms,
heterotrophs, which acquire the free energy to fuel their structural effects
from objects with a rest mass where energy is stored in their chemical bonds.
Plants, by contrast, are autotrophs, which acquire energy directly from the
sun by absorbing photons.
Basic
kinds of animals. Since animal behavior guidance systems evolve at
each level, the most basic divisions among animals correspond to the levels
of part-whole complexity in the biological behavior guidance system discussed
in the last section. We should expect some kinds of animals at each level
of biological organization, because animals are just a variety of the living
objects that evolved at each stage of evolution. They are, in technical terms,
heterotrophs, which acquire the free energy to fuel their structural effects
from objects with a rest mass where energy is stored in their chemical bonds.
Plants, by contrast, are autotrophs, which acquire energy directly from the
sun by absorbing photons.
At the prokaryotic level of biological organization, the heterotrophs are bacteria. They feed on energy-rich molecules, and their varieties mirror the kinds of molecules they consume.
The animal behavior guidance system at this level need not be very complex. It is a molecular structure in the cell wall that can identify the right kinds of molecules and draw them through the cell wall into the cell to be dismantled inside while keeping dangerous (and useless) molecules outside.
To be sure, autotrophs at this level, such as cyanobacteria, also consume molecules, but only for use as parts in growing (or as input signals), not as a source of free energy. Parts can be acquired without an animal behavior guidance system, because the simplest, energy-poor molecules, which are almost always available as part of the medium both inside and outside the cell and pose no danger, can be used as parts. But animals need a special system in order to ingest large, energy rich molecules, for that poses a hazard.
At the eukaryotic level of biological organization, the heterotrophs are animal-like protists, or single-celled eukaryotes. They feed mainly on prokaryotes and plant-like protists (algae). But there are several new animal behavior guidance systems, and to suggest their range, let me mention three kinds.
The paramecium has a stiffened cell membrane with an oral groove along one side in which many hair-like cilia sweep small particles into a mouth, where they are enclosed in a vacuole for digestion inside the cell — unless they are harmful and they are kept out. This animal behavior guidance system is clearly independent of the biological behavior guidance system, the nucleus, for when the nucleus is removed, a paramecium continues moving and feeding for many days.
Amoebas have a soft plasma membrane, instead of the stiffened cell membrane that gives paramecia their shape, and amoebas move along the substratum by constructing an extension of the plasma membranes on one side of the body while dismantling it on the other (pseudopodia). Amoebas ingest prokaryotes, algae, and even small animal like protists by wrapping part of their outer membrane around the energy-rich object and then enclosing the package within its cytoplasm as a food vacuole where digestion takes place. Hence, their choice about feeding is made locally in the course of moving about.
Finally, collar flagellates remain attached to the substratum and use the whip-like motion of their flagellum to pull microorganisms to a region near a collar at the base of the flagellum where they can be ingested.
There is one kind of protist, Euglena, whose identity as plant or animal is in doubt, because these flagellates have chloroplasts for acquiring energy from photons and yet also ingest prokaryotes. However, they may be plants, rather than animals, because they probably use their prokaryotic prey for their nutrients, rather than for energy, since what is scarce near the surface of the ocean which they inhabit are heavier atoms. (This is the kind of scarcity that accounts for animal-like multicellular flowers in rain forests, such as the Venus fly-trap, that ingest animals. But since the objects are ingested for parts, rather than energy, they are not animals by this definition.)
The multicellular level of biological organization makes possible the most powerful ways of guiding animal behavior, because it uses multiple neurons. Neurons are cells whose structure enables them to transmit signals from one place in the body to another or among themselves, and since neurons are capable of being organized at a series of levels of part-whole complexity, there are differences among kinds of multicellular animals that are nearly as basic as these differences among other kinds of animals.
Multicellular animals direct behavior at other objects by the contraction of muscle cells located where the muscles are needed to change the body’s shape, and neurons generate behavior by controlling the relevant muscles. But behavior must depend on input from the other objects in space in order for animals to choose how to behave towards them and to generate behavior whose spatial aspects will control the outcome of interacting with them, and so neurons must be connected to sensory receptors on which other objects in space have an effect. The choice of kind of behavior depends on the “interneurons” connecting sensory neurons on the input side and motor neurons on the output side (usually in conjunction with input about the state of the body itself concerning its energy needs, mating urges, damage to cells, and the like).
The behavior of the neuron itself is generated, as in all machines, by the geometrical structure of such material objects and how they structure the flow of free energy toward entropy increase in their region. In the neuron, molecular mechanisms in the plasma membrane control of the passage of sodium and potassium ions through its plasma membrane in such a way that an action potential propagates along the axon of the neuron, and that affects the passage of small proteins, called “neurotransmitters,” at the synapses where they meet determines whether a signal is passed on. But the details of the neuron's mechanism are well known and not relevant here.
What is relevant is how they constitute an animal behavior guidance system. Any behavior guidance system, given its function of selecting between incompatible kinds of behavior, must have systems serving three basic sub-functions, and in those that are made of neurons, they will be called the sensory input system, the goal selection system and the behavioral output system.
Among animals with nervous systems, the most basic classification has to do with the level of part-whole complexity in the systems serving the three basic sub-functions. The animal behavior guidance system imposes a geometrical order on the region by how it moves its body around and acts on other objects in space, and there are higher levels of neurological organization in multicellular animals, because they are possible and each actualizes further the power that animals can have by taking spatial aspects of the objects on which they act into account in guiding their behavior in relation to them. The levels of part-whole complexity occur in the three basic systems making up the animal behavior guidance system, and given the ontological explanation of revolutionary evolution, they lead, as we shall see below, to three additional stages of evolution in multicellular animals. Somatosensory animals are the first animals to evolve at the multicellular level, and three additional stages occur because of levels of neurological organization: telesensory (most invertebrates and non-mammalian vertebrates), subjective (mammals), and manipulative (primates).
At the social level of biological organization, there are two basically different kinds of animals, only one of which has a new kind of animal behavior guidance system that is possible only at the social level. That is the spiritual animal, which is made up of primates as its lower level organism. The same behavior guidance system serves as both its biological and animal behavior guidance system, because the behavior of many individual primates is coordinated so that the spiritual animal acts as a whole by the use of language. The capacity to use language depends on the evolution of additional levels of neurological organization, which determines a series of stages of evolution (though, as we shall see, the last level does not require any change in the biologically inherited structure of the nervous system).
Primitive spiritual animals are groups of primates with the capacity to use a language of natural sentences (that is, sentences with a simple subject-predicate grammar). The linguistic level of neurological organization comes from a higher level of part-whole complexity within the primate faculty of imagination in the so-called “left brain” which enables it to combine the meanings of words as propositions and to indicate the structure of the covert linguistic act involved by verbal behavior (by words and grammatical markers).
Rational spiritual animals are also animals at the social level of biological organization, but they have the capacity to use psychological sentences (that is, sentences with predicates formed of verbs of propositional attitude, such as “believes” and “desires,” together with complete sentences). This linguistic capacity enables the users of language able to think about the causes of their behavior and belief as part of the process of causing them, and so it will called the reflective level of neurological organization. Reflection makes it possible for language to guide individual level and social level behavior in a new way, which involves rational imagination and enables them to understand arguments, and that is the foundation for a new form of evolution by reproductive causation, namely, cultural evolution.
Philosophical spiritual animals are social level animals in which culture has evolved a new level of argument, in which all the arguments of the rational level are organized as parts of a single argument. That is a means of proving that propositions are necessarily true, and thus, it is a new way of discovering the true and the good. Though this is a stage of cultural evolution, it gives the members of the spiritual animals in which it exists a higher level of rational imagination, and thus, it is also the philosophical level of neurological organization. But there are, as we have seen, two ways of constructing such philosophical arguments, epistemological and ontological, and only one of them is able to succeed.
This classification of animals identifies most of the natural kinds, but there is another kind of animal it leaves out. It may be represented by the other kind of animal at the social level of biological organization, or multisomatic animals, such as insect colonies. They have long posed a puzzle for zoologists, because it is not obvious how to classify them. They are social and, thus, like human society, and yet they are so different. But insect colonies are only one of a number of animals that have defied classification, and this ontological explanation of evolution, including both gradual evolution and revolutionary evolution, offers a solution. Thus, we may use the way in which it solve the problem of anomalous animals as a further reason to believe that it is true, before we take up the structure of the nervous system and explain how it works in detail.
 Anomalous
Kinds of Animals. There is another way that the animal
behavior guidance system combines with the levels of biological organization
to account for species as natural kinds. Thus far, animals have been classified
according to the new kind of animal behavior guidance system made possible
by each biological level — as bacteria, animal-like protists, and animals
with nervous systems, and spiritual animals. But the same elements can be
used to explain the various anomalous kinds of animals.
Anomalous
Kinds of Animals. There is another way that the animal
behavior guidance system combines with the levels of biological organization
to account for species as natural kinds. Thus far, animals have been classified
according to the new kind of animal behavior guidance system made possible
by each biological level — as bacteria, animal-like protists, and animals
with nervous systems, and spiritual animals. But the same elements can be
used to explain the various anomalous kinds of animals.
It requires only that we notice that organisms with the new kind of animal behavior guidance system made possible by one biological level can also be organized merely as so many lower level organisms bundled together as a higher level animal organism. Though it does not have the new kind of animal behavior guidance system made possible by its own level of biological organization, it is still an animal, because it is a heterotroph. Its only animal behavior guidance system is the kind that evolved at the lower level of biological organization. And it has many of them, one in each of its parts.
The most obvious example is the sponge. Sponges are multicellular organisms. And they are animals, acquiring energy from energy-rich objects. But unlike other multicellular animals, they lack a nervous system. The reason is that the sponge is just an organization of collar flagellates, each with an animal behavior guidance system of a kind that evolved in collar flagellates on the eukaryotic level, but put together as a multicellular organism. These animal .like protists are configured like a jar; their flagellums pull water through pores in its side and push it out an opening at the top. Each cell feeds on microorganisms that gather around the collar at the base of its flagellum, and they share their energy with other kinds of cells that hold them together and form pores to the outside. However, the basic choice required by their animal nature is made by the individual cells. The sponge has no animal behavior guidance system as a whole. It would be just a multicellular plant, except that its cells are animal-like rather than plant-like. (See diagram of anomalous animals.)

The cellular slime mold is a similar kind of animal, except that the parts are amoeboid animal-like cells, rather than collar flagellates. When a spore from the cellular slime mold lands where food is available, it feeds, grows and reproduces repeatedly, spreading out as far as food is available. But when food runs out, they all move back toward the center, climb on top of one another and form a slug-like animal, which may move around as a unit for a while. Eventually, however, it becomes a stalked, fruiting body from which new spores are released to try their luck at finding new feeding areas. The biological behavior guidance system coordinates their behavior, using messenger proteins to determine the cells to have different kinds of behavior. Their bodies become polarized front to back; each secretes messenger molecules from one end and moves toward the same molecules that are secreted by others. However, the choice required by its animal nature, between feeding or not on the objects it encounters, is made by the animal behavior guidance systems of its amoeboid members. (See diagram of anomalous animals.)
Fungi are so anomalous that they are almost always classified as a third kingdom, separate from both plants and animals. But the recognition that the only animal behavior guidance system that some animals have is an animal behavior guidance system located in their parts that evolved at a lower level of biological organization reveals that fungi are animals of an anomalous, but natural kind.
Fungi are basically eukaryotes, because they have a nucleus. But their animal behavior guidance system is at the prokaryotic level. Fungi acquire energy by ingesting energy-rich molecules (usually after secreting special proteins to promote decomposition outside their bodies), and thus, they need only the prokaryote's mechanisms for selecting which kinds of molecules to admit and which kinds to exclude. Many such mechanisms are located in their cell wall.
In fact, this prokaryotic animal behavior guidance system is organized on two higher levels of biological organization. There are fungus-like protists, such as yeast, at the level of the single-celled eukaryote, and there are fungi, like mushrooms, at the multi-cellular level. (Multicellular fungi are actually multi-nucleated organisms, because their many nuclei inhabit an interconnected labyrinth of cell walls containing many prokaryotic animal behavior guidance systems operated jointly. A multi . nucleated structure is possible, because fungi have no mitochondria to keep under control. But the reason that they lack mitochondria is probably that the prokaryotes from which fungi acquired their animal behavior guidance system did not use the oxygen-consuming process to extract energy from energy-rich molecules.)
There are, therefore, three levels of animals with the simplest kind of animal behavior guidance system: bacteria, single-celled fungi, and multicellular fungi (see diagram of anomalous kinds of animals).
Finally, this same way of combining animal behavior guidance systems with levels of biological organization accounts for what might be called “multisomatic animals,” including insect colonies, hydrozoa (colonies of hydra), corals, and the colonies of blind mole-rats in East Africa. In each case, the only animal behavior guidance systems are the nervous systems contained in the multicellular animals of which the multisomatic organism is composed. Thus, the organism's behavior as a whole depends mainly on the biological behavior guidance system at the multicellular level, and it reproduces as a whole. (See diagram of anomalous animals.)
In insect colonies, for example, the mating flight of the queen sets up a whole new colony composed of her offspring. But sexual reproduction involves the construction of a entire new colony from a fertilized egg cell (at the eukaryotic level). Thus, it must construct organisms at two levels of part-whole complexity. After constructing the queen’s body, her body is used to construct the other members, and the queen becomes the center of a multisomatic structure in which pheromones are used to coordinate the behavior of her multicellular offspring (the same mechanism used to coordinate cells at the multicellular level).
The limitations of such a mechanism are worth noting. It must make sure that pheromones are received by the right animal parts, but since its multicellular parts do nor remain attached to one another like cells in a multicellular organism, the biological behavior guidance system must require the multicellular animals to have certain unchanging spatial relations, or else it could not use the locations of the pheromones to give them different kinds of behavior. Thus, multisomatic organisms have hives, labyrinths of tunnels, or the like in which pheromones can be distributed, and though its parts have locomotion, the whole multisomatic organism is basically sedentary.
All the kinds of animals, including these anomalous heterotrophs, fall naturally into one or another of the categories generated by combining the concept of an animal behavior guidance system with the concept of levels of biological organization. Animals that lack the new kind of animal behavior guidance system that evolves at their own level of biological organization are made of lower level organisms each with the animal behavior guidance system that evolved at a lower level of biological organization.
The classification of insect colonies as an anomalous kind of animal at the social level does suggest, however, that a new kind of animal behavior guidance system may be possible at the social level of biological organization. That is the possibility realized that is realized in a unique way by the evolution of language among manipulative animals, that is, as spiritual animals.