![]()
![]()
 2.
Prokaryotic stage: DNA molecules. What evolves at the second stage
are DNA molecules, which are a higher level of biological organization than
RNA molecules, and the major accomplishment of this stage is the living cell,
currently represented by bacteria and other prokaryotic cells.
2.
Prokaryotic stage: DNA molecules. What evolves at the second stage
are DNA molecules, which are a higher level of biological organization than
RNA molecules, and the major accomplishment of this stage is the living cell,
currently represented by bacteria and other prokaryotic cells.
In general, a new stage of evolution is caused ontologically by a higher level of part-whole complexity in the organisms that have already evolved. But as we have seen, in order to be a higher level organism, the lower level organisms must be bundled together as structural causes in a way that coordinates their behavior both in doing the work of reproducing itself and in doing the non-reproductive work that controls conditions affecting its reproduction.
An evolutionary stage in which organisms with a higher level of part-whole complexity evolves gradually in the direction of natural perfection for organisms of its kind is inevitable, if it is both possible and functional (that is, the higher level of organization can be tried out as a random variation and it is able to control an entire range of conditions affecting reproduction that were out of reach during the previous stage).
 The
inevitability of DNA proto-organisms. In this case, the lower level
organisms are simpler proto-organisms, the RNA molecules that evolved at the
first stage of evolution, though as we have seen, different varieties of RNA
were to some extent combined in favorable regions where they all shared the
benefits of the useful proteins being synthesized by any of them. And the
upper level organism that begins the second stage of evolution is itself merely
a higher level proto-organism, at least initially, for it starts off as just
a way of attaching RNA-like molecules to one another as parts of a bigger
macromolecule: the DNA molecule. DNA proto-organisms can, therefore, be the
ontological cause of another stage of evolution, and a second evolutionary
stage is inevitable, because such a higher level of biological organization
is both possible and increases the power to control relevant conditions.
The
inevitability of DNA proto-organisms. In this case, the lower level
organisms are simpler proto-organisms, the RNA molecules that evolved at the
first stage of evolution, though as we have seen, different varieties of RNA
were to some extent combined in favorable regions where they all shared the
benefits of the useful proteins being synthesized by any of them. And the
upper level organism that begins the second stage of evolution is itself merely
a higher level proto-organism, at least initially, for it starts off as just
a way of attaching RNA-like molecules to one another as parts of a bigger
macromolecule: the DNA molecule. DNA proto-organisms can, therefore, be the
ontological cause of another stage of evolution, and a second evolutionary
stage is inevitable, because such a higher level of biological organization
is both possible and increases the power to control relevant conditions.
 The
possibility of DNA. There is no special problem about showing how it
was possible for the evolved organisms to try out the radical variation that
would begin the second stage, because gradual evolution during the first stage
had already coordinated the behavior of RNA molecules in doing both kinds
of work and useful combinations of varieties of RNA were already combined
in favorable locales.
The
possibility of DNA. There is no special problem about showing how it
was possible for the evolved organisms to try out the radical variation that
would begin the second stage, because gradual evolution during the first stage
had already coordinated the behavior of RNA molecules in doing both kinds
of work and useful combinations of varieties of RNA were already combined
in favorable locales.
The DNA molecule, like RNA molecules, is a strand of nucleic acids linked by a sugar-and-phosphate backbone, but the kind of sugar used in DNA enables complementary strands of its nucleic acids to bind together and wind up as a double helix, which is a more stable molecule than strands of RNA. There is no reason to doubt that the range of random variations on the process of RNA reproduction was wide enough to try out DNA molecules made up of copies of evolved RNA molecules.
The random variations included the substitution of a new kind of nucleotide substituted for one of the (four) kinds used in RNA and a new kind of sugar backbone for holding members of the new set of nucleotides together as a DNA molecule. The single strand of DNA could then become a double helix by synthesizing its own complement, that is, by each nucleotide attracting its complement and binding them together as a second stand of DNA.
In order to go through reproductive cycles, organisms must be able to generate both essential kinds of structural effects, reproduction and various forms of non-reproductive work. That was possible in this case, because the various structural causes bundled together in of DNA molecules were copies of lower level proto-organisms that were able to do both kinds of essential work at the previous stage.
The DNA molecule could do non-reproductive work, because it contains multiple parts, usually called "genes", each of which has a geometrical structure that can serve as a template from which the RNA molecule (called messenger-RNA, or mRNA) can be transcribed. And DNA molecules could synthesize proteins in the established way, because the ribosomes and tRNA needed for protein synthesis had already evolved and were readily available in colonies of RNA in every local region. To be sure, transcription is a structural effect that requires the help of special protein catalysts. But natural selection was so inefficient at the first stage that they would be among the proteins being synthesized in at least some colonies of RNA. And a template for such proteins would be among the first genes included in DNA molecules, since their expressions would control a condition that affects their reproduction.
DNA reproduction also requires a special protein to promote it, but such a protein could be supplied in a similar way at first. And with the addition of one kind of nucleotide to those available in such colonies, DNA molecules would have all the parts they need to reproduce themselves in much the same way as RNA..
At the beginning, DNA molecules would be able to reproduce and control conditions affecting their reproduction, because there were favorable conditions. Colonies of RNA would provide most of the energy, parts, ribosomes and other housekeeping molecules, and DNA molecules themselves could direct the synthesis of the new protein molecules needed to control the condition affecting their own reproduction.
That is, by containing a gene whose protein facilitated transcription of mRNA and another gene whose protein facilitated its own replication, DNA molecules would themselves enrich the mix of molecules in the region sufficiently to go through reproductive cycles alongside RNA in the same regions, reproducing themselves and doing non-reproductive work during the same period of the circadian cycle as RNA.
The second level of biological organization illustrates a feature typical of biological levels: not only is the organism going through reproductive cycles on a higher level of part-whole complexity in space than the organisms from the previous stage, but each reproductive cycle of the higher level organism usually involves many reproductive cycles of lower level organisms. That is, in the biological levels, the reproductive cycles at the next stage have a higher level of part-whole complexity in time as well as a higher level of part-whole complexity in space.
DNA molecules generate non-reproductive structural effects by transcribing segments as mRNA molecules and using the mRNA molecules to synthesize a protein molecule, and this process may occur many times from the same gene.
 The
greater power of DNA. Nor is it difficult to see why this higher level
of part-whole complexity in evolving organisms would be able to control conditions
affecting reproduction beyond what was possible at the previous stage. The
favorable environment provided by the gradual evolution of RNA toward natural
perfection for proto-organisms of their kind is what made it possible for
DNA molecules to go through cycles in which their parts (genes) did the non-reproductive
work of synthesizing protein and they reproduced themselves as a whole. And
DNA would evolve by reproductive causation, because they were also driven
through reproductive cycles by the cycle of night and day, along with RNA.
The
greater power of DNA. Nor is it difficult to see why this higher level
of part-whole complexity in evolving organisms would be able to control conditions
affecting reproduction beyond what was possible at the previous stage. The
favorable environment provided by the gradual evolution of RNA toward natural
perfection for proto-organisms of their kind is what made it possible for
DNA molecules to go through cycles in which their parts (genes) did the non-reproductive
work of synthesizing protein and they reproduced themselves as a whole. And
DNA would evolve by reproductive causation, because they were also driven
through reproductive cycles by the cycle of night and day, along with RNA.
The main source of the greater power of DNA to control additional relevant conditions is their higher level of part-whole complexity. They have traits with a higher level of part-whole complexity. They could generate many non-reproductive structural effects by directing the synthesis of various proteins, and thus, they could bring conditions under control that required special combinations of proteins. Some of the powers they evolved are described below.
Moreover, their higher level traits would evolve faster than colonies of RNA, because useful combinations of proteins would be selected more reliably by the success of a DNA molecule in reproducing as a whole than they were at the first stage. (Combinations of proteins were selected at the first stage by the success of the several independent RNA templates for those proteins being more likely to populate favorable regions when storms came because of their greater population.)
For example, when storms redistributed both RNA and DNA molecules, not enough of the RNA molecules may arrive at the same locale for any of them to be of much use. But DNA molecules with a useful combination of genes could direct the synthesis of a number of unusual proteins at a single location, and both DNA and RNA would be able to complete reproductive cycles.
Furthermore, the range of random variations from which a natural selection was made was likewise greater, because in addition to new genes, DNA molecules could try out new combinations of established genes (especially if, among the kinds of proteins that had evolved by then for interacting with DNA, there were some that could cut DNA molecules and link them together in different ways).
It is inevitable, therefore, that proto-organisms with a higher level of biological organization would begin a second stage of evolution. We know from our ontological explanation that, if reproductive cycles generated by higher level organisms are both possible and more powerful, they are inevitable and another stage of evolution will inevitably begin. And since, as we have seen, DNA molecules satisfy those conditions, they would begin a second stage of gradual evolution.
There may be other ways of combining RNA molecules as parts of a higher level proto-organism which could also go through reproductive cycles. But our ontological explanation of evolution as a global regularity implies that the most powerful among them becomes the foundation of subsequent evolution, and thus, we know from the structures of extant organism that DNA molecules were the most powerful.
 Gradual
evolution of DNA. Without tracing its course in detail, we can
sketch some of the more significant accomplishments of the gradual change
of DNA molecules in the direction of maximum holistic power for proto-organisms
of its kind, and that will put us in a position to explain the origin of life.
Gradual
evolution of DNA. Without tracing its course in detail, we can
sketch some of the more significant accomplishments of the gradual change
of DNA molecules in the direction of maximum holistic power for proto-organisms
of its kind, and that will put us in a position to explain the origin of life.
 Regulatory
mechanism. As we have seen, its higher
level of biological organization enabled DNA to do non-reproductive
work requiring the cooperation of multiple protein molecules. But that was
only the first step in the increase of power that was possible for this higher
level organism, because power could increase with better coordination of the
behavior of the RNA-level proto-organisms.
Regulatory
mechanism. As we have seen, its higher
level of biological organization enabled DNA to do non-reproductive
work requiring the cooperation of multiple protein molecules. But that was
only the first step in the increase of power that was possible for this higher
level organism, because power could increase with better coordination of the
behavior of the RNA-level proto-organisms.
There is some coordination in the synthesis of proteins in the fact that their templates, or genes, are all parts of the same DNA molecule. That would make DNA molecules more powerful than RNA molecules, even if it was a random process that selected which genes on the DNA to use in synthesizing proteins. But DNA molecules could evolve far more powerful ways of coordinating the behavior of the lower level organisms, for there are mechanisms that can make the synthesis of special sets of proteins depend on the conditions that prevailed in the local colonies of RNA they encountered.
The geometrical structure of DNA would make such a random variation possible. When complementary strands of DNA wind up and form a double helix, the genes they contain cannot do either essential kind of work. Thus, if the DNA molecule was normally kept closed during the phase of the cycle when its structural causes were doing non-reproductive work (except, perhaps, for a certain region of genes whose proteins facilitated mRNA transcription, DNA replication, or universal housekeeping functions), it could serve as a mechanism for coordinating the non-reproductive behavior of the lower level organisms of which it is composed. It required only the evolution of genes whose proteins would respond to molecules in the region indicating that a certain condition prevailed by opening up certain segments of DNA, allowing the genes located there to be transcribed into RNA for protein synthesis, and closing them up again when the conditions changed.
The evolution of proteins that promote, repress, or de-repress segments of DNA would also enable DNA to generate discriminative responses to situations, controlling only the conditions that needed to be controlled. DNA molecules with such a regulatory mechanism could succeed in reproducing, while DNA molecules without it failed, because they would control relevant conditions more efficiently, that is, doing more with less.
The evolution of such a regulatory mechanism would enable DNA to respond to each situation in which it found itself by coordinating the behavior of its parts to control the conditions that happened to be relevant there. But that did not exhaust the new powers that were possible with a regulatory mechanism, because it also became possible to give the behavior of lower level organisms a temporal structure.
Once DNA had evolved the power to synthesize different combinations of proteins in response to different situations, the selection of rather minor random variations would make it possible for a single DNA to act on the world in a way that involved a series such coordinated responses. Messenger molecules signaling that some condition had been brought under control by one behavior could be the trigger for initiating another kind of behavior to control the next. The ability to make one change in the environment and then another when the first was completed would bring new relevant conditions under control.
Non-reproductive structural effects with a temporal structure could evolve for controlling new relevant conditions even though other relevant conditions in the region were being controlled more or less jointly (with RNA and its "housekeeping" proteins playing a supportive role to a variety of DNA in the region). It would make a difference locally which would tend to affect the reproduction of the DNA that generated the behavior, because it would be located nearby. But it would also enable its colony to be selected over others at the level of the group when storms or other disturbance redistributed all the molecules.
 The
cell. The power of this DNA-based regulatory system to coordinate the
behavior of its parts as a sequence of responses to changing conditions was
not fully exploited, however, until the evolution of a cell wall which enclosed
all the molecular interactions generated by a single DNA molecule and isolated
them from the surrounding colony. The cell, perhaps, marks the point at which
proto-organisms evolve into something must be called an organism. The cell
wall made it possible for complex sequences of behavior to add up reliably
over time to a definite local result.
The
cell. The power of this DNA-based regulatory system to coordinate the
behavior of its parts as a sequence of responses to changing conditions was
not fully exploited, however, until the evolution of a cell wall which enclosed
all the molecular interactions generated by a single DNA molecule and isolated
them from the surrounding colony. The cell, perhaps, marks the point at which
proto-organisms evolve into something must be called an organism. The cell
wall made it possible for complex sequences of behavior to add up reliably
over time to a definite local result.
Cell-wall barriers may have evolved from the protein-based mechanism whose function was to keep regulatory proteins attached to DNA molecules after reproducing, for the reproduction of the whole DNA-based regulatory mechanism involved not only copying the DNA molecule itself, but also supplying new regulatory proteins for the copy made. This is consistent with prokaryotes, in which the loop of DNA remains attached to the cell wall.
Whatever their original function, however, once barriers became a wall separating the regulatory mechanism and its structural effects on molecules from the rest of the world, DNA molecules would be able to do non-reproductive work by generating behavior that involved complex sequences of RNA-level responses. The change made by each step in such a sequence could build on the effect of the previous step, and thus, the behavior of the cell as a whole could control conditions that were out of reach for naked DNA molecules.
There was, of course, a price to be paid for adding a cell wall to the DNA-based regulatory system. The cells were part of the structure that had to be reproduced along with the DNA molecules in order to reproduce the whole higher level organism. Thus, DNA would have to generate non-reproductive behavior that would construct a new cell wall during one phase, and it would have to divide the cell as part of its reproductive behavior during the other phase. But these are precisely the kinds of conditions that could be controlled by DNA with the ability to generate various combinations of proteins in sequences over time with cumulative local effects.
To be sure, once it was enclosed in a cell, DNA could not rely on RNA in the region to supply the “housekeeping” proteins, but had to include the genes that supply them. By the same token, however, this assured the natural selection of such DNA, for they could overcome the recurrent scarcity of parts and energy in RNA colonies by tapping sources of free energy in territories that were uninhabitable by RNA.
The evolution of cell walls once again increased the efficiency of reproductive causation. When the organisms going through cycles of reproduction were simply proto-organisms depending on the molecules available in a local colony to use free energy to do work, like RNA molecules, they were selected for synthesizing functional proteins by being located where their reproduction was more likely to be affected by them and by being reproduced in such numbers that they were more likely to wind up in favorable, local regions after a storm. Though reproductive causation does not need any more than a weak connection between useful non-reproductive work and reproduction, its efficiency is increased when natural selection is more discriminating.
DNA took the first step toward coordinating the behavior of RNA molecules by tying the fates of various genes together as parts of a single molecule, but, without a cell, the control of relevant conditions was still a joint project in each region, with different kinds of DNA benefiting from one anothers secondary effects and RNA playing a supporting role to all the DNA molecules located there by supplying the "housekeeping" proteins.
But once there was a cell to contain all the proteins synthesized by a single DNA, all the conditions required to go through a reproductive cycle could be controlled by a single organism. This concentrated the global reproductive cycle into many local reproductive cycles, and the cell as a whole became gradually more powerful in controlling all the conditions that affected its reproduction. (See diagram of reproductive cycle of nonliving organism.)
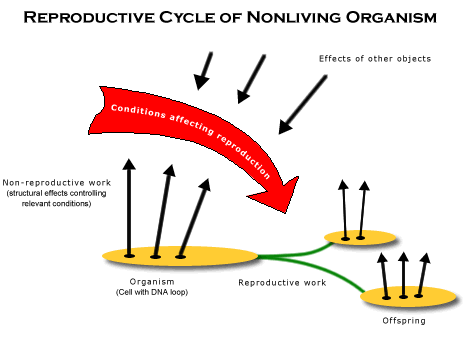
A DNA-based regulatory mechanism contained by a cell is the kind of complex material structure whose reproductive cycles constitute, along with space, the basic reproductive global regularity. It can go through reproductive cycles as a whole, because it can do both kinds of essential work. It coordinates the behavior of its genes to do non-reproductive work by transcribing mRNA whose protein products control relevant conditions, such as acquiring energy and parts from the environment and constructing more cell wall. It also coordinates the behavior of its genes to do reproductive work by transcribing mRNA whose proteins prepare for DNA replication, cell division, and carry out the sequence of changes involved in reproducing. But there is one further step that would make such cells even more powerful in controlling relevant conditions.
 The
Biological Behavior Guidance System. Even
with the evolution of a cell wall, organisms based on DNA are still
driven through reproductive cycles by the cyclic change in the supply of free
energy due to the rotation of the planet. But this simple organism would be
more powerful, if it could go through reproductive cycles on its own, independently
of the circadian cycle.
The
Biological Behavior Guidance System. Even
with the evolution of a cell wall, organisms based on DNA are still
driven through reproductive cycles by the cyclic change in the supply of free
energy due to the rotation of the planet. But this simple organism would be
more powerful, if it could go through reproductive cycles on its own, independently
of the circadian cycle.
Such cells would be more powerful because they would be able to go through reproductive cycles faster than organisms still entrained by the cycle of night and day, thereby consuming the free energy and parts before others could. They could also tap energy sources in locations that were previously uninhabitable because they were not affected in the right way by the circadian cycle.
In order to complete reproductive cycles on its own, however, it needed a system for selecting which of the two essential kinds of behavior to generate at any moment and for generating it, because DNA cannot transcribe mRNA and replicate itself at the same time.
Such a system required only a relatively minor change. Since the DNA-based regulatory mechanism could already express different genes depending on the kinds of messenger molecules present at any moment, the presence of a messenger molecule indicating that the cell had gathered enough energy and grown large enough could trigger the switch to reproduction. And it could switch back to growth again when another kind of molecule, which was present only when reproduction was complete, opened up the growth-genes. But since the chemical interactions involved in doing both kinds of work evolved under the energy conditions imposed by the cycle of night and day, it may also have required other new protein mechanisms to adapt them to the new conditions. (For example, to synthesize proteins at night, it may have had to replace the free energy formerly provided by high energy photons).
Let us call this mechanism for coordinating behavior the "biological behavior guidance system." Though some of its functions have been served by DNA from the beginning, all the steps we have traced in its gradual change in the direction of greater power must be traversed before DNA has accumulated the additional material structures required to be the ontological cause of all the structural global regularities needed to go through a cycle of reproduction on its own (when the region contains free energy that it can tap and other resources, such as the parts it needs for doing both kinds of work).
A higher level of part-whole complexity in organisms going through reproductive cycles is what causes a new stage of gradual evolution, and that requires the higher level organism to have some way of coordinating the behavior of the lower level organisms. That was done implicitly by DNA from the beginning by combining many genes, the equivalent of RNA proto-organisms, as parts of a single complex molecule, and we have traced how reproductive causation gradually increased the complexity of the higher level proto-organism, adding regulatory proteins and a cell wall, so that the higher level organisms could, as a structural cause, generate more complex behavior as a whole. Finally, the behavior it generates as a whole comes to include selecting between growth and reproduction and generating each of them when selected. At that point in the second stage of evolution, it serves all the functions of a biological behavior guidance system.
Though behavior guidance is a form of non-reproductive work that controls reproduction, that does not erase the difference between reproductive and non-reproductive work. An organism is still a bundle of structural causes, and in addition to coordinating all their behavior, it must be able to reproduce its structure as a whole. Reproduction is still unique: it is the construction of a material object with a geometrical structure like itself. And reproductive work requires parts throughout the whole prokaryotic organism to move and interact with one another in a fundamentally different way from non-reproductive work. Besides copying its DNA molecule and synthesizing regulatory proteins for its offspring, it must divide its cell by dismantling it in some places and constructing new cell wall in a different way from simply growing. That requires the DNA molecule and its regulatory mechanism to move and interact in basically different ways from doing non-reproductive work. What is different when the biological behavior guidance system evolves is that selecting when to reproduce becomes one of the kinds of behavior generated as part of its non-reproductive work, that is, a structural global regularity involving the transcription of mRNA and synthesis of proteins. That means it must set up the conditions that will not only result in its reproduction, but also put the biological behavior guidance system back in a condition in which it can select and generate other kinds of behavior when it is done reproducing. Thus, when the non-reproductive work of the cell comes to include selecting between phases of growth and reproduction, that does not make reproduction a form of non-reproductive work, but merely controls when reproduction occurs.
With the evolution of a biological behavior guidance system in the DNA-containing cell, what exists are organism of the same kind as bacteria, which are called “prokaryotes” or “prokaryotic cells.”
The prokaryotic biological behavior guidance system is the locus of changes in subsequent evolution. Those evolutionary changes are all basically kinds of non-reproductive work, or behavior, which the prokaryotic cell uses to control additional conditions that affect its reproduction, and since the biological behavior guidance system is an aspect of the geometrical structure of the whole organism that can determine whether it grows or reproduces, relatively minor modifications in that structure can select among and generate additional kinds of behavior. It is based on the regulatory mechanism, whose function was to generate various kinds of complex structural effects, but when it becomes part of a biological behavior guidance system, as aspect of the structure of the whole prokaryotic cell has the function of selecting and generating all its behavior. Thus, the evolution of new kinds of behavior involve changes in the behavior guidance system.
 The
origin and nature of life. Having show how the cell-enclosed DNA-based
regulatory system evolves into a full-fledged biological behavior guidance
system, we have derived from spatiomaterialism a material structure of the
kind that is ordinarily said to alive. No one doubts that prokaryotic cells,
such a bacteria, are living organisms. That would mean that in a spatiomaterial
world like ours, the evolution of life is inevitable. It is one of the powers
that organisms acquire during the second stage of evolution.
The
origin and nature of life. Having show how the cell-enclosed DNA-based
regulatory system evolves into a full-fledged biological behavior guidance
system, we have derived from spatiomaterialism a material structure of the
kind that is ordinarily said to alive. No one doubts that prokaryotic cells,
such a bacteria, are living organisms. That would mean that in a spatiomaterial
world like ours, the evolution of life is inevitable. It is one of the powers
that organisms acquire during the second stage of evolution.
Life does not begin, therefore, deep in the ocean where heat is vented from the center of the Earth. It cannot, because there is no cycle in the supply of free energy, like the circadian cycle near the surface. It only appears to be where life began, because that is the ecological niche to which prokaryotes adapted that has changed least since the origin of life, and so it contains some of the earliest models of prokaryotic cells (called "archebacteria").
It is worth noting, however, what this explanation of its origin implies about the nature of life, for it suggests that life has an essential nature. To say that life has an essence is to say that there is a real difference — not just in our minds, but in the nature of things — between living and non-living organisms. Our ordinary understanding of life can tell us where to look for the difference, but in order to be sure what the nature of life is, those intuitions must be confirmed by an ontological cause. Life is a product of reproductive causation, but there should be some moment between the lifeless molecules with which evolution begins and the complex living organisms to which it leads, when a change occurs that is so basic that we recognize it as what our intuitions were seeing only confusedly.
Our ordinary conception of the nature of life seems to be captured by the notion of autonomous activity, given that it occurs naturally, rather than as just an artifact designed by beings like us. By "activity", I mean, not just a disposition, or physical property of the kind mentioned in the last chapter, but rather a more complex kind of goal-directed behavior in which different kinds of dispositions are generated in concert and feedback from these consequences is used to bring about some kind of overall change. And by “autonomy,” I mean that different kinds of activities are determined, not merely as responses to conditions that arise, as if they were guided by feedback in attaining some further goal, but rather internally, by something that selects among them. These are the traits to which we would point in order to remove any doubts that more highly evolved organisms are alive, and if we survey the significant steps in gradual evolution toward greater power, the step at which life begins may stand out.
We called RNA molecules “proto-organisms,” because they are already complex material structures that can do both essential kinds of work. (Each nucleotide attracts its complement in reproducing itself, and each triplet attracts a certain kind of amino acid in the non-reproductive work of protein synthesis.) Though they are on a higher level of part-whole complexity than most ordinary molecules, those ways of doing work are just dispositions, that is, mere physical properties. They are hardly “activities,” and there is nothing “autonomous” about them, since they are entrained by the circadian cycle.
The DNA molecule’s higher level of part-whole complexity enables it to coordinate the behavior of many RNA-level proto-organisms in doing both kinds of work. But neither kind of structural effect involves goal-directed behavior. Though its behavior as a whole may involve the synthesis of various protein molecules, they do not use feedback from the consequences of previous behavior to generate the kind of behavior that brings about some goal. It is still not activity in the relevant sense.
With the evolution of a regulatory mechanism, however, the DNA's higher level of organization does make the use of feedback possible. DNA can respond to different situations in different ways, and thus, the whole complex surely coordinate the behavior of many RNA-level organisms in one way in response to the consequences of coordinating the behavior of other RNA-level organisms in some other way. Though the use of feedback to attain the same goal in different ways in different situations makes it goal-directed, it is not yet enclosed in a cell wall, and so it is still just a series of responses that is largely controlled by the environment. Its activity lacks autonomy.
Enclosing the DNA-based regulatory mechanism within a cell makes its goal-directed behavior more autonomous, because it is able to coordinate the behavior of many lower level proto-organisms as a sequence of responses within its cell, each of which may depend on feedback from the last, and thereby carry out a temporally complex activity. That is what justified calling it an “organism,” rather than a mere “proto-organism.” But its activity is not fully autonomous, because its goals are given. Though such behavior may pursue different goals in different situations, the cell is not fully autonomous as long as its goal depends on the situation. Since its goals are still just variable means to a more encompassing, constant goal, the cell does not seem to alive. It is arguably just reacting to changes in its environment in ways that attain some overall goal, which is not itself chosen.
This final objection is overcome, however, when the cell-enclosed DNA-based regulatory mechanism evolves into a biological behavior guidance system, because at that point, its goals are not all dictated by changes in the environment. What determines when to grow and when to reproduce is no longer the circadian cycle, but the cell itself. It selects which goal to pursue for itself, depending on its own condition, mainly whether it has grown sufficiently large. Growth and reproduction are both goal-directed activities, but they are not simply sub-goals pursued as means to some constant overall goal. They are the two basically different kinds of work that an organism must do in order to evolve by reproductive causation.
Reproduction is not the goal of non-reproductive work, as we have seen, but rather the cause of evolutionary change. What changes is mainly the non-reproductive work. The organism acquires ways of behaving that make it increasingly powerful at controlling conditions that affect its reproduction, and the control of such a condition is the goal of every non-reproductive structural effect. What makes non-reproductive work functional is contributing to the holistic power of the organism. Thus, its function is not reproduction. Though reproduction may be a goal-directed activity, it is merely the kind of activity that causes population growth, scarcity and natural selection and does not itself have a function.
Furthermore, the power to choose between growth and reproduction on its own is, as we have seen, a form of non-reproductive work. It is another condition that is controlled by its behavior as a whole, that is, by coordinating the behavior the organisms of which it is composed. The cell-enclosed DNA-based regulatory mechanism opens up special segments of its DNA for transcription into mRNA and translation into proteins, while closing other, in order to set up the process by which it reproduces. That is the original function of the biological behavior guidance system.
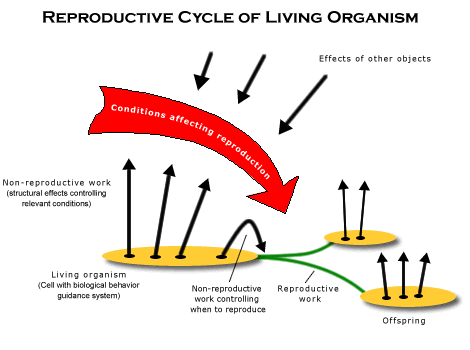
The reason for calling this mechanism a “behavior guidance system” is that its function is not merely that of a mere cybernetic system, which uses feedback to adjust behavior in order attain some constant goal, but selecting between incompatible kinds of behavior. In the case of the biological behavior guidance system, it is selecting between phases of growth and reproduction, which cannot be generated at the same time. (There is, as we shall see, an additional kind of choice between goals that must be made in animals, and thus, the mechanism that serves that function will be called the “animal behavior guidance system.”) The function of selecting between incompatible kinds of behavior requires the behavior guidance system to have a structure that also serves to two other functions: input and output. It must have input from appropriate conditions in the world (including conditions in the organism itself) in order to select the right kind of behavior, and it must be able to generate one or another kind of behavior as output. In all, therefore, three functions are involved in behavior guidance, one involving choice, another involving input for making that choice, and a third for generating the behavior chosen as output. It is the output of a behavior guidance system that corresponds to a cybernetic system, because generating behavior often involves the use of feedback to guide it toward the goal that has been chosen. Although feedback also requires a kind of input, it does not necessarily use the same input mechanisms as the those that serve the function of choosing, for these input have different functions.
Judging by what is ordinarily meant by “life,” therefore, life begins when reproductive causation produces organisms with a biological behavior guidance system, for at that point, they can decide for themselves when to switch from pursuing one of at least two incompatible goal-directed activities. Though choosing between these goals still depends on input indicating when it has grown large enough or when reproduction is complete, it is autonomous in the sense that it is no longer merely responding to its environment as an external master. These cells are able to go through cycles of reproduction on their own, independently of the cycle of night and day. Life begins, therefore, when reproductive causation makes organisms powerful enough to remove themselves from the natural mold supplied by the large scale structure of the universe, the circadian cycle, and go through reproductive cycles on their own. Life begins with the autonomy of selecting between the two basic activities involved in reproductive cycles.
This concept of the essence of life implies that viruses and plasmids are not alive. They do not have a way of selecting between their two kinds of mechanical effects, but rather depend on living objects around them for their reproduction, just as primary structures did before the evolution of life.
This intuition about the essential nature of life is confirmed by its ontological significance, for the autonomy of the cell's activity marks a dramatic change in the ontological cause of evolution. We called organisms going through reproductive cycles the ontological cause of gradual evolutionary change, because given the wholeness of space, their motion and interaction had to add up over time to a global regularity in which their non-reproductive work becomes increasingly powerful at controlling relevant conditions. But such reproductive cycles are just a derivative ontological cause, since they are constituted by more basic ontological causes. And until the evolution of the biological behavior guidance system, the ontological causes constituting reproductive cycles were of two radically different kinds: the organism itself as structural cause generating both kinds of work and the large scale structure of the universe. The latter supplied not only the free energy required for the structural cause to do work, but also the temporal structure of the reproductive cycle, the cycle of night and day. However, there was a radical change in the ontological cause of evolutionary change when life evolved. Reproductive cycles become the autonomous activity of material structures. That is, the structural cause took over responsibility for the temporal structure of the reproductive cycle, leaving the large scale structure of the universe merely the role of supplying the free energy to be consumed in doing work. Thus, except for how reproductive cycles still depend on a supply of free energy, the beginning of life meant that the ultimate ontological cause from which the ontological cause of evolution derives is a complex material structure.
This dramatic change in the ontological cause of evolution means that, from this point on, evolution is something that organisms do to themselves. What causes natural selection, according to reproductive causation, is reproduction itself, because the multiplication of organisms in the region causes scarcity. As long as proto-organisms or organisms were driven through cycles of reproduction by the cycle of night and day, natural selection could be seen as something that was being done to them. But when the biological behavior guidance system evolved, structural causes not only do both essential kinds of work, but also generate the temporal structure of the cycle as a whole. Thus, evolution becomes something that living organisms do to themselves. Evolution still depends, of course, on the free energy supplied by its star. But given such a thermodynamic flow of matter from potential energy through kinetic energy and photons toward evenly distributed, life itself is what makes life evolve. That seems to be ontological confirmation enough of our intuition that life is essentially the kind autonomous activity that begins with the evolution of the biological behavior guidance system.
![]()