![]()
![]() Reproductive
causation: Gradual evolution. The first of the two main kinds of
reproductive global regularities is gradual evolution, and it will be explained
here by considering, first, its Ontological cause,
the existence of reproductive cycles in closed or isolated regions
of space, and then, its Ontological
effect, a global regularity in which evolutionary change leads gradually
in the direction of an optimum, which
will be called “natural perfection” because it makes the most of what is possible
in a spatiomaterial world. In other words, reproductive causation implies
that gradual evolution is progressive.
Reproductive
causation: Gradual evolution. The first of the two main kinds of
reproductive global regularities is gradual evolution, and it will be explained
here by considering, first, its Ontological cause,
the existence of reproductive cycles in closed or isolated regions
of space, and then, its Ontological
effect, a global regularity in which evolutionary change leads gradually
in the direction of an optimum, which
will be called “natural perfection” because it makes the most of what is possible
in a spatiomaterial world. In other words, reproductive causation implies
that gradual evolution is progressive.
Though I have already suggested why gradual evolutionary change is ontologically necessary (in the introductory section of Reproductive global regularities), there is more to be said about its ontological cause. First, in order to show that reproductive cycles are an ontological cause, it is necessary to show how they derive from space, matter, and the ontological causes of simpler global regularities. Second, in order to explain what gradual evolution involves, it is necessary to explain what is meant by “natural perfection” and to show how this ontological cause generates gradual change in the direction of natural perfection in both the organisms and the ecology.
![]() Ontological
cause: reproductive cycles. What
constitutes the ontological cause of gradual evolution is a way that all the
simpler ontological causes of global regularities work together. Their combination
yields a kind of (derivative) substance with an odd nature. It is the reproductive
cycle. It is basically just an organism with heritable traits that normally
reproduces itself during its lifetime. But explaining the derivative substances
ontologically will show the necessity and character of the global regularities
they cause.
Ontological
cause: reproductive cycles. What
constitutes the ontological cause of gradual evolution is a way that all the
simpler ontological causes of global regularities work together. Their combination
yields a kind of (derivative) substance with an odd nature. It is the reproductive
cycle. It is basically just an organism with heritable traits that normally
reproduces itself during its lifetime. But explaining the derivative substances
ontologically will show the necessity and character of the global regularities
they cause.
Reproductive cycles are, in effect, “substances” that derive from material and structural causation. Enplaning that will show how reproductive cycles are reduced to spatiomaterialism, since we have already seen how those more elementary ontological causes are reduced to spatiomaterialism..
Material global regularities are involved, because what is going on in reproductive cycles is the use of free energy to do work, that is, to make things happen that would not otherwise happen. As we shall see, most of the relevant free energy is supplied in the way sketched earlier, that is, by the radiation from a star. That is a thermodynamic flow of matter toward evenly distributed heat, and when it is intercepted by a planet in orbit around the star, it can be used to fuel reproductive cycles. Cycles of reproduction are irreversible processes, though each reproductive cycle involves many structural global regularities.
Structural global regularities are involved because what uses the free energy to do work are structural causes. There are material structures that coincide with the thermodynamic flow of matter toward evenly distributed heat, and the ensuing motion and interaction results in certain specific effects, which are events that would not otherwise occur. This is an ontological explanation of how efficient causes produce their effects, as we have seen in the case of dispositions. In discussing structural global regularities, we saw that structural global regularities entail efficient-cause connections. What makes such structuring of the tendencies to kinetic energy and randomness into a new ontological cause is how structural causes are combined in the constitution of reproductive cycles.
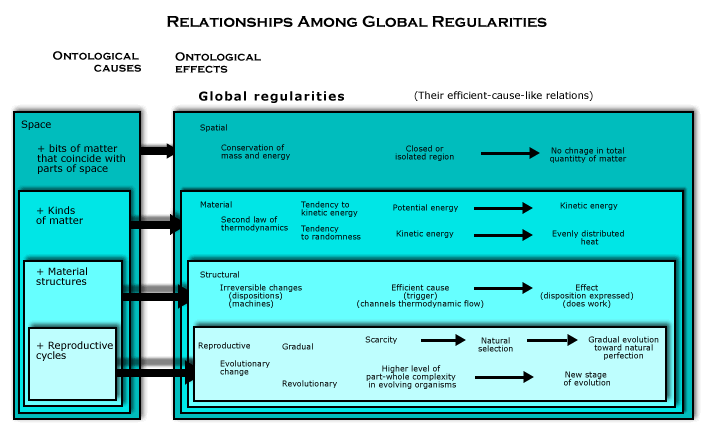
Reproductive cycles. What makes the reproductive cycle a new ontological cause is the kind of material structure that generates this structural global regularity (which is ordinarily called an "organism"). It is a kind of material structure that generates a cycle of irreversible processes that includes both reproduction and non-reproductive work.
The organism. The organism is a complex material structure, for it includes a number of material structures bundled together as various structural causes. But in addition to the distinctive structural global regularity that each such a structural cause generates, there is a structural global regularity in which the material structure as a whole is reproduced, making the whole process generated a reproductive cycle.
Such material structures will be called "organisms," or "reproducing organisms, when they go through reproductive cycles on their own. But after a few stages of evolution, as we shall see, there may also be a few levels of part-whole complexity of such material structures within the reproducing organism, and so it will be useful to refer to material structures of this kind as "primary structures." That is, primary structures are complex material structures that are able to reproduce themselves, and the use of this technical term will make it easier to describe the structures of organisms at later stages of evolution.
Organisms (and primary structures) generate, therefore, two kinds of structural global regularities during a reproductive cycle. Each structural cause that is bundled together in the organism usually generates a different kind of structural global regularity. But the organism can also generate a structural global regularity as a whole in which it reproduces itself as a whole, including all the structural causes bundled together in it. In order to keep these two radically kinds of ontological effects of its structural causes straight, I will call the former “non-reproductive work” and the latter “reproductive work.”
I will call the structural global regularities generated by structural causes "structural effects" in order to have a simpler way of referring to them.
In organisms (and primary structures generally), many structural causes are bundled together, and since the structural global regularities that each generates involves a connection between an efficient cause and its effect, we can also say that different efficient-cause connections are bundled together in the organism. But that means that the structural cause is not just an ontological cause, but also an efficient cause, for its inclusion in the structure of the organism is sufficient to bring about the effect during the reproductive cycle. Such an effect is simply the difference that the structural cause makes to what tends to happen (in the organism, its behavior, or in the environment as a result of its behavior) because of the structural global regularity it generates. (Or as the technical formula for our ordinary notion of efficient cause goes, it is a non-redundant member of the set of conditions in the organism that is sufficient for the effect.) Such an effect is what I will mean by "structural effect."
Talk of “structural effects” ignores the difference between the material structure as the ontological cause of the structural global regularity and the inclusion of the material structure in the organism as an efficient cause (along with other conditions) of its occurrence during the reproductive cycle. But that is useful in describing organisms, because different parts of the organism are responsible for different structural global regularities and, thus, different structural effects.
The difference between the reproductive work and non-reproductive work of structural causes can, therefore, be put in these terms. Since the structural effect is the work done by the structural cause in generating a structural global regularity, the two radically different kinds work that are essential to organisms (and primary structures) can also be described as the organism's “reproductive structural effect” and its “non-reproductive structural effects.”
This will be useful, because in all organisms (and primary structures), including the most basic, it is not only the organism as a whole that has both a reproductive and non-reproductive structural effect. Each of the structural causes bundled together in the organism has both a reproductive and non-reproductive structural effect. In fact, the reproductive structural effects of the parts is what enables the organism to reproduce as a whole, for it is a process in which each part reproduces itself.
The non-reproductive structural effects of the material structures bundled together in an organism (or primary structure) are often called the "traits" of the organism. But notice that when traits are explained ontologically, their heritability by offspring is entailed. Since traits are non-reproductive structural effects of the material structures bundled together in organisms, they are necessarily inherited by offspring, because such parts of organisms (and primary structures) also have reproductive structural effects as part of the reproductive work by which the organism as a whole reproduces.
The cyclic structural global regularity. In order for reproductive cycles to exist, however, organisms must actually go through cycles in which they do both their reproductive and non-reproductive work. It is not enough for their material structures to endure though time without doing both kinds of work.
When structural causes are bundled together spatially as an organism, a number of different non-reproductive structural effects are bundled together. But these structural effects do not necessarily occur at the same time. The various structural causes often (though not always) have their non-reproductive structural effects at different times. But to hold that the organism goes through a cycle is to hold that its parts have structural effects that ultimately put the organism back in much the same position where it began, so that it can go through another cycle. That means that the bundle of structural effects has a temporal structure as a whole. Thus, not only are the structural causes bundled together in space as a complex physical organism, but their structural effects are combined in time as a cycle. It is a cycle of changes.
What makes the cycle a reproductive cycle, however, is that at some point during the cycle, the organism is reproduced. That is, one or more additional organisms of the same kind are constructed and put in a position to go through reproductive cycles side by side. The reproduction of the organism as a whole is essential to reproductive cycles being a new kind of derivative ontological cause.
In all the basic organisms, as we shall see, reproduction depends on each structural cause bundled together in them having a reproductive structural effect as well as a non-reproductive structural effect. (This is rather straightforward in the case of bacteria and protists, but it also holds in a more complicated way for multicellular organisms, as we shall see.)
Notice that crystals do not go through reproductive cycles of this kind. The reason crystals are sometimes said to go through reproductive cycles is that, as each molecule is added to the growing crystal, the structure required for the addition of another molecule is created, enabling the processes to be repeated. But this is not the kind of reproductive cycle that is an ontological cause of reproductive global regularities, because the crystal is not an organism (or primary structure) by our definition. It does not have a non-reproductive structural effect distinct from its reproductive effect, not to mention that with just one structural cause, it cannot be a bundle of structural causes at all.
Natural selection. Reproductive global regularities are caused ontologically by space and reproductive cycles (as a derivative ontological cause), because reproductive cycles add up in space over time to natural selection. That is, as they endure through time in the region, they reproduce, and since their own population increase makes free energy (or other resources) scarce, they impose natural selection on themselves.
Reproductive cycles endure through time like substances because they are cyclic. Each cycle begins a new cycle, and thus, the cycles have a continued existence.
Unlike basic substances, however, derivative substances can be destroyed. But since there is both a spatial and a temporal aspect to reproductive cycles, there are two ways that this kind of derivative substance can cease to exist. A cycle can go out of existence either because the material structure of the organism going through the reproductive cycle is destroyed, or because the organism is unable to generate its entire cycle of structural effects. But as long as they do not go out of existence, they are like substances, enduring through time.
As reproductive cycles, however, they also multiply in space. Endurance through time also involves, in the case of reproductive cycles, the reproduction of the organism going through them. That is, in addition to reproducing in time as one cycle follows another, they also reproduce in space as additional organisms of the same kind are constructed.
Reproductive cycles would not be a new kind of (derivative) ontological cause, if all they did was endure through time, for then they would be a mere cyclic irreversible structural global regularity, like a motor. The reproduction of the organism is what makes them a new derivative ontological cause.
Space is the other ontological cause of evolution, along with reproductive cycles.
Spatial causation is the only simpler kind of global regularity that has not been mentioned thus far in explaining the ontological cause of evolution. But spatial causation is one of the ontological causes of every global regularity, because the wholeness of space is what makes the motion and interaction of bits of matter in a region add up over time. And adding up in space to natural selection is just another such role for spatial causation.
In causing material and structural global regularities, however, the structure of space within the region was relevant in another way as well. Its geometrical structure was needed to explain why potential energy tends to become kinetic energy and why kinetic energy tends to become randomly distributed heat. And structural global regularities depend on the geometrical structure of space not only because it helps constitute material structures (that is, material objects with a geometrical structure), but also because it enables them to move around in space without changing their geometrical structures.
In the case of gradual evolution, by contrast, the geometrical structure of space within the region is not directly relevant. It is only the wholeness of space that works together with cycles of reproduction to constitute reproductive global regularities. Thus, the role of space as an ontological cause of evolution depends only on the fact that all the parts of space in the region fit together uniquely as a whole, much like how it worked together with matter to cause the conservation of matter.
Being contained by space makes reproductive cycles add up in a unique way as time passes. Since the continued existence of reproductive cycles entails the multiplication of reproductive cycles in space, expanding the population of organisms, it combines with the space that contains them to cause ontologically a scarcity that imposes natural selection on them.
Though the total matter in any isolated region of space does not change, there is a way that push comes to shove when some of the matter constitutes reproductive cycles. As the organisms going through them reproduce themselves, generation after generation, the reproductive cycles not only endure through time, but also multiply in space, and thus, the population of organisms grows. But the matter in the closed or isolated region must add up arithmetically in space as time passes, that is, as a total that does not change over time (according to the principle of the conservation of matter). Though some of the matter in the region must exist in the form of free energy in order for material structures to do work of any kind, there is only a finite thermodynamic flow of matter from potential energy through kinetic energy (and photons) to evenly distributed heat in any isolated region of space, such as a planetary system or the surface of a planet. Thus, as reproductive cycles multiply in space, scarcity of free energy, if nothing else, will eventually limit the number of reproductive cycles that can continue to exist in the region. Some reproductive cycles will have to come to an end.
In other words, organisms going through cycles of reproduction impose natural selection on themselves. That is how the wholeness of space works together with reproductive cycles to generate a new kind of global regularity. It is simply how reproductive cycles add up in space as time passes.
To be sure, it is natural selection, and not just random selection, because the organisms are not only reproducing, but also doing non-reproductive work which can control conditions that affect reproduction. Those differences among organism are, as we shall see, why natural selection leads to a gradual change in the direction of natural perfection. What makes such variations among organisms possible is that organisms have non-reproductive as well as reproductive structural effects. But what forces a selection to be made at all is their reproduction in space as well as time.
Random variations. This explains all but one of the three elements generally assumed to be part of Darwin’s mechanism. Since heritable variations and natural selection are entailed by the nature of reproductive cycles as an ontological cause, only random variations remain to be explained. There must also be random variation in the organisms in order for natural selection to cause evolutionary change. But random variations are due to another fact also implicit in the nature of reproductive cycles as a ontological cause.
What makes variations in the traits of organisms possible is the fact that organisms (and primary structures) are complex material structures, that is, that they are bundles of various kinds of structural causes. If different varieties of structural causes can be bundled together as a complex material structure at all, they can be bundled together in different ways. That is, different bundles can include different kinds of structural causes. New structural causes can be added, both new kinds and additional particular material structures. And the same structural causes can even be bundled together in different ways. The part-whole complexity inherent in structural causes being bundled together as organisms makes an enormous range of kinds of reproductive cycles possible.
The possibility of random variations is actualized in the simplest organisms, because the global regularity caused ontologically by their material structures is merely a tendency. The structural global regularity that they generate includes the construction of imperfect copies as well as perfect copies of their material structures, thereby introducing random variations.
Material structures have their ontological effects by how they channel the thermodynamic flow of matter toward evenly distributed heat. It depends on how the geometrical structures of the material objects involved coincide with the geometrical structures inherent in the thermodynamic flow of matter toward evenly distributed heat (the geometrical structure of potential energy being converted into the kinetic energy of objects and the geometrical structure of the nonrandom distribution of their causally relevant factors becoming random). Thus, when various material structures are involved (such as an organism and the many parts used to construct copies of it) or free energy is supplied to them in various ways (for example, by radioactive decay as well as chemical energy and the usual photons), what is ontologically necessary about the future may be only a tendency. In particular, the structural global regularities generated by simpler organisms include the construction of imperfect copies as well as perfect copies of their complex material structures. The imperfect copies will tend to resemble the structure of the organism being reproduced, but there will be random variations on it.
This is not to say that structural global regularities, as global regularities, are inherently probabilistic. With the addition of other material structures, it is possible to ensure that no errors occur in reproduction, as in some higher organisms. Reproduction is imperfect in simpler organisms because their material structures cannot control all the structural causes that may be relevant to what happens. The structural global regularity includes various different outcomes with different probabilities. In higher organisms, where reproduction is infallible, special mechanisms are required to introduce random variations.
Darwin’s mechanism as a consequence. Thus, the way that reproductive cycles add up over time in the wholeness of space includes all three of the elements generally considered essential to Darwin’s mechanism (natural selection, heritable traits, and random variations).
The bundles of structural causes that generate reproductive cycles as structural global regularities make up the populations of reproducing organisms in which Darwin’s mechanism is assumed to be at work.
Natural selection is caused by how reproductive cycles add up in space over time, that is, by the scarcity, due to population growth, that requires some reproductive cycles to come to an end.
Traits are inherited by offspring, because the traits are the non-reproductive structural effects of the structural causes bundled together as organisms and those same structures are what is copied in reproduction.
And there are random variations on the traits of organisms, because reproduction is a structural global regularity that involves only a tendency to make perfect copies (or there are mechanisms for introducing random variations).
All three elements of Darwin’s mechanism are entailed in the ontological cause of evolution as a reproductive global regularity.
It may seem paradoxical or even perverse to take reproductive cycles, rather than reproducing organisms, to be the ontological cause that helps constitute the reproductive global regularities. Organisms are three dimensional objects, which endure through time like other substances, whereas reproductive cycles are four dimensional objects, which endure through time only in the sense that the changes involved are cyclic and can go indefinitely. And since reproductive cycles are derivative substances, constituted by reproducing organisms (and a thermodynamic flow of matter toward evenly distributed heat), reproducing organisms do work together with space to cause ontologically the same reproductive global regularities.
There are two reasons for preferring to think of reproductive cycles as the ontological cause of evolution (along with space). The first is that reproductive global regularities are fundamentally different from simple structural global regularities, and the source of that difference is that reproduction occurs cycle after cycle (for it is the scarcity due to their population increase that imposes natural selection on them). And since an ontological cause is something that endures through time like a substance (or a basic relationship among substances), what causes evolution ontologically is the continued existence of reproductive cycles, not just the continued existence of reproducing organisms. What endure is a series of four dimensional objects with the property of multiplying in space.
The other reason has to do with the nature of the ontological effect. What changes in the entire region, as we shall see, is the kinds of reproductive cycles. Though these cycles are implicit in the organisms that are their ontological causes, the nature of changes that occur in evolution has to do with how they fit into the structure of the whole cycle, and we keep that in mind, by recognizing that the causally relevant unit is the whole cycle, rather than just the three dimensional organism.
Since reproductive causation entails all the essential elements of Darwin’s mechanism, it can explain everything currently explained in evolutionary biology. But this ontological derivation of them is not merely proof that there is an ontological necessity about what has already been discovered by empirical science. It is the beginning of a proof of new propositions about the course of evolution, first of all, that evolution is a gradual change in the direction of natural perfection.
![]() Ontological
effect: gradual change toward natural perfection. Reproductive
causation implies that evolution is progressive. The reason for calling it
progressive is that the global regularity constituted by reproductive cycles
and space involves change in the direction of natural perfection. That is,
as we shall see, how reproductive cycles add up in space over time. But this
is also to introduce the foundation for explaining the nature of goodness,
which is one of the mortgages that must be paid back in order to use spatiomaterialism
as an ontological foundation, and thus, it is appropriate to start by explaining
what I mean by “natural perfection” and making clear how it affords an explanation
of the nature of goodness.
Ontological
effect: gradual change toward natural perfection. Reproductive
causation implies that evolution is progressive. The reason for calling it
progressive is that the global regularity constituted by reproductive cycles
and space involves change in the direction of natural perfection. That is,
as we shall see, how reproductive cycles add up in space over time. But this
is also to introduce the foundation for explaining the nature of goodness,
which is one of the mortgages that must be paid back in order to use spatiomaterialism
as an ontological foundation, and thus, it is appropriate to start by explaining
what I mean by “natural perfection” and making clear how it affords an explanation
of the nature of goodness.
Natural perfection. A dictionary would tell us that “perfect” refers, in general, to the most complete, whole, or mature state of a thing, that is, a state without defects, a state in which no change would make it superior in any relevant way. Things can be perfect in different ways, but in every case, what makes the perfect stand out is that it is an optimal state that can hold in a certain kind of part-whole relation. And what I mean by “natural perfection” is a certain kind of optimal part-whole that is most appropriate to the basic nature of what exists in a world like ours, where everything is constituted by matter and space enduring through time.
What is perfect is always a whole that is made up of parts in some way. Though there are many different ways that parts can make up whole, the whole must be more than just the sum of its parts. There must be relations among the parts. And to be optimal, its parts must be of the right kinds and numbers and they must be related in the right ways. Thus, in general, a part-whole relation can be said to be optimal when it makes the most out of the least in some salient, determinate way. Though it depends on the kind of part-whole relation involved, perfection is an optimum that comes from combing the fewest and simplest parts of some kind so that the whole makes the most of them in a salient way.
Something can be said to be perfect of its kind, therefore, to the extent that it makes the most of things of its kind in the appropriate way. Beautiful works of art might be said to be perfect of their kind, because, given a suitable, classical definition of beauty, they involve such an optimal part-whole relation. But what I mean by “natural perfection” involves a far more basic part-whole relation, because perfection of its kind is a part-whole relation that makes the most of what exists in a natural world, that is, of the substances that endure through time.
In a spatiomaterial world like ours, perfection would have to make the most of space and matter enduring through time. Though its basic nature is just a form of matter in motion, it is not fair to paint the portrait of the world as a bleak picture of cosmic indifference. To living things, at least, there seems to be a difference between good an bad. And the significance of the law of entropy increase is not, despite its universality, that the fate of the world is a heat death in which everything has the same temperature. Rather, its significance is that there is such a thing as free energy, for that affords a salient way in which the most can be made of the parts in a spatiomaterial world like ours. Though entropy never decreases, the large scale structure of the universe is such that potential energy is constantly becoming evenly distributed heat, and since that means that there is a constant supply of free energy, there is a salient way that the most can be made out of what exists. As we have seen, material structures can use free energy to do work, and if such structural causes were to use as much of the free energy available in the region to do the work of controlling as much as possible what happens in the world, such a world would obviously have an optimal part-whole relation of the kind by which “perfection” is defined.
An optimal part-whole relation is one that makes the most out of the least, and in the case of something that is naturally perfect, the whole does the most in the sense of exercising maximum power, given the available free energy, and it does so with the least in the sense that it uses the fewest and simplest structural causes needed to exercise that power. That is an optimum that is often called “maximum efficiency,” though as we shall see, that does not characterize what is involved in natural perfection adequately.
Maximum efficiency is the kind of optimal part-whole relation that is the goal in designing machines. In the case of automobiles, for example, the goal is maximum power, reliability and efficiency in serving its function (say, safe transportation) at the least cost in parts, effort of construction and energy consumption. And since machines eventually wear out, the goal is a machine in which all the parts wear out at the same time.
Natural perfection involves maximum efficiency, but it is a more basic and, thus, more general kind of optimal part-whole relation, because it also involves doing as much as possible in the way of using free energy to control what happens in the world. The ultimate natural perfection is a optimum in which any relevant change in the parts or their relationships would make the whole less powerful.
Natural perfection is the direction of evolution by reproductive causation, but as we shall see, natural perfection involves several part-whole relations that are optimal in this sense. Two of them characterize the direction of gradual evolutionary change, the natural perfection of organisms and the ecology, as we shall see shortly. But gradual evolution is only one of the global regularities generated by reproductive causation, for there are also revolutionary episodes in evolution. Together, as we shall see, these two global regularities entail change in the direction of a natural perfection with an overall structure on planets (or in planetary systems) like ours.
Goodness. It should be noted at the outset that natural perfection plays a very important role in this ontological argument, because it affords an explanation of the nature of goodness, enabling us to pay back one of the four mortgages we took out in order to use spatiomaterialism as the foundation for proving these necessary truths in the first place (that is, along with an explanation of consciousness, how Einsteinian relativity could be true, and the existence of something worthy of worship).
Goodness can be defined in terms of natural perfection, because perfection is a kind of part-whole relation. Natural perfection is the property of the whole in such optimal part-whole relations, and so goodness can be defined as the property of the parts. That is, goodness can be explained ontologically as the property of contributing to the natural perfection of the whole of which it is part. And since all forms of natural perfection fit together in a necessary way as part of the overall structure of natural perfection, there are no ultimate conflicts about what is good, and goodness is simply the property of contributing to natural perfection.
Every part of the optimal part-whole relation that defines natural perfection is good, but since the perfect whole does the most with the least in the way of parts, each part makes a unique contribution. No part is redundant (though many parts of the same kind may be needed for the whole to be perfect).
By this definition of “good,” therefore, what is good ought to exist, because it is “called for” by natural perfection. That is, if anything that is good did not exist, the whole would not be naturally perfect. There would be a change that would make it more perfect. That is the sense in which natural perfection can be said to "call for" its existence.
Goodness is just a property of parts in relation to natural perfection, but as we shall see, it explains why all the things that are ordinarily considered to be good are good, including goals that are good for beings like us. It even explains an the aspect of goodness that makes such a theory seem impossible, namely, why rational beings ought to choose the good. And it reveals goals to be good that are not generally recognized as such today.
If evolution is progressive in the sense of involving change in the direction of natural perfection, it is also progressive in the sense of making things good (or bad). Progressive evolution is, in other words, the source of goodness in a spatiomaterial world, and since such evolutionary change is inevitable in a spatiomaterial world like ours, the ultimate source of goodness is the nature of the world itself.
The reason for believing that evolution by reproductive causation is progressive is that it is change in the direction of natural perfection, and in the end, there will be at least five different ways in which evolution by reproductive causation makes the world naturally perfect. They all fit together as part of a necessary overall structure, and since there are no basic conflicts to cast doubt on the whole being naturally perfect, it will be clear that the change involved in reproductive global regularities is indeed progressive. The first global regularity caused ontologically by space and reproductive cycles is gradual evolution, and it is change in the direction of the natural perfection of organisms and the natural perfection of the ecology. And gradual evolutionary change is itself a kind of natural perfection. Thus, three forms of natural perfection will be discussed below. Two further forms of natural perfection will be introduced later in this argument (both having to do with stages of evolution).
The natural perfection of organisms is maximum holistic power. The parts of this optimal part-whole relation are the structural causes that are bundled together as an organism going through reproductive cycles. Its power is holistic, because it depends on all the non-reproductive work done by its structural causes and all those parts must be of the right kinds and combined in the right ways. Its power is maximized in the sense that those non-reproductive structural effects control as many of the conditions affecting its reproduction as possible for organisms of its kind. But since free energy is consumed in generating structural effects, power is also maximized when the fewest and simplest structural causes are used because free energy is used most efficiently. Since that is to do the most with the least, maximum holistic power is a form of natural perfection, which will be called the “natural perfection of organisms.”
By the ecology, I mean how organisms exist alongside one another in regions of space. There must be an ecology, because organisms reproduce in space and go through reproductive cycles alongside one another. And there is a natural perfection about the ecology that complements the natural perfection of the organisms, because there is an optimal part-whole relation at the ecological level as well. It is also a kind of maximum holistic power. The parts, in this case, are the organisms going through reproductive cycles, and the whole is a power that depends on all the organisms in the region. Again, the whole is not merely the sum of the parts, because it depends on the kinds of organisms in the region, the numbers of each, and how they interact. In this case, however, holistic power is measured by how much of the free energy available in the region is being used by all of them to fuel their reproductive cycles. Given the kind of perfection that is appropriate in a spatiomaterial world like ours, that is the most salient way of doing the most that can be done. And as we shall see, it is also a case of doing the most with the least, because the organisms that use the free energy are each maximally efficient. I will call it the “natural perfection of the ecology.”
There is even a natural perfection about the process of gradual evolution itself, for there is a way in which evolution also makes the most out of the least when it is due to reproductive causation. In this case, the parts are all the moments in the course of evolution, and the whole is the overall course of evolution itself. Evolution by reproductive causation makes the most out of each moment, because the moments all add up as time passes to change in the direction of the natural perfection of organisms and their ecology. That is, since evolution is progressive, it is itself a form of natural perfection, which will be called the “natural perfection of evolutionary change.”
In order to show that evolution by reproductive causation is progressive, I will show that reproductive cycles add up in space as time passes to a gradual change in the direction of natural perfection. The first step is to see why organisms change gradually in the direction of maximum holistic power, and then why the ecology evolves gradually in the direction of their maximum consumption of the free energy available in the region.
![]() Gradual
evolution of the organism. The new derivative ontological cause
that has been derived from spatiomaterialism is the reproductive cycle, and
reproductive cycles add up in space as time passes by imposing natural selection
on themselves. Reproduction is a cause of natural selection, because resources
are finite and population growth eventually makes free energy (if not other
resources) in any isolated region scarce. Though such scarcity is a change
in the environment of each organism, it is not caused by anything external.
It is internal, since reproduction is an aspect of reproductive cycles. And
since population growth eventually leads to scarcity in a world like ours,
some organisms will eventually be unable to survive and reproduce. But as
long as there are new, heritable traits within the range of the traits being
“tried out” by random variations that would make organisms better able to
control the conditions affecting their reproduction, natural selection would
tend to favor individuals with new traits that increase their power. Their
greater reproductive success would eventually change the population until
every member had the new trait (that is, acquired the new power).
Gradual
evolution of the organism. The new derivative ontological cause
that has been derived from spatiomaterialism is the reproductive cycle, and
reproductive cycles add up in space as time passes by imposing natural selection
on themselves. Reproduction is a cause of natural selection, because resources
are finite and population growth eventually makes free energy (if not other
resources) in any isolated region scarce. Though such scarcity is a change
in the environment of each organism, it is not caused by anything external.
It is internal, since reproduction is an aspect of reproductive cycles. And
since population growth eventually leads to scarcity in a world like ours,
some organisms will eventually be unable to survive and reproduce. But as
long as there are new, heritable traits within the range of the traits being
“tried out” by random variations that would make organisms better able to
control the conditions affecting their reproduction, natural selection would
tend to favor individuals with new traits that increase their power. Their
greater reproductive success would eventually change the population until
every member had the new trait (that is, acquired the new power).
If that is how reproductive cycles add up in space as time passes, we can see how evolutionary change would lead to the natural perfection of the organisms going through them. Reproduction would be the main cause of evolution, if these conditions held of organisms that start off simple, uniform and weak and it were possible for new traits to be tried out and bundled together with others in the organism, because evolutionary change of this sort would go on for a long time, adding one new trait after another, making the organisms complex, diverse and powerful. Indeed, it needn’t stop until the organisms are as powerful as possible for organisms of their kind, that is, until there are no more new power-enhancing traits within the range being tried out by random variations on existing organisms. That would be their maximum holistic power, because at that point, no change in the causal connections bundled together in their reproductive cycles that is possible for organisms of their kind would make them any more powerful.
This is to restrict the range of possible traits to those that can be "tried out" by random variations during their gradual evolution, and that depends on the nature of the organism and how random variations are generated (that is, the nature of the structural global regularity). It may be possible to imagine useful traits that fall outside that range, but that would not show that the organisms had a less than maximum holistic power in the relevant sense.
Example of amphibian evolution. To illustrate how reproductive causation could generate such a regularity, consider how it would explain fish evolving into amphibians. Reproduction among fish of various kinds would eventually cause a scarcity in the usable energy and other resources available in the water. Those fishes in which random variations happened to try out traits that enabled them to move their bodies across land, would find plants and other animals that provided a new source of usable energy. Those fish would tend to succeed in reproducing, while otherwise they might have failed.
Something like this apparently occurred among lungfishes about 345 million years ago. These fresh water fish had already evolved lungs, perhaps to absorb oxygen from the air when the water was stagnant and deficient in oxygen, and they had “lobe fins,” or large fleshy bases for their paired pectoral and pelvic fins, making it possible for them to move across land. At first, they were relatively uniform, simple and barely able to complete cycles of reproduction requiring locomotion across land. But reproductive causation would make them increasingly complex, diverse, and powerful. As random variations on their inherited multicellular structure tried out new traits, one new trait after another would be added. Each new trait would make them more powerful at controlling conditions that affected their reproduction, and as their complexity and power increased, new ways of controlling conditions could be tried out by random variations. Thus, fins would gradually evolve into legs, enabling them to crawl more efficiently across land, and since land plants and insects provided many different sources of usable energy, the different ways of acquiring energy would lead to the evolution of different species of amphibians. Amphibians would go on adding one new trait after another, increasing their power, until none of the new traits that could be tried out by random variations on their multicellular structures could make them better able to control conditions that affect their reproduction. Evolutionary change would stop only when they were as powerful as possible for organisms of their kind in whatever environment they occupied.
Such a proliferation of species is called a “radiation,” and it would occur again and again. When a more basic random variation on amphibians finally tried out internal fertilization (instead of fertilization in the water), making it possible for eggs that remained on land to house a form of embryological development that did not involve a larval stage, reptiles would be able to acquire usable energy from new sources on land. Two legged locomotion would be the start that gave dinosaurs their day in the sun. Those with wing-like limbs would begin a radiation of birds.
Long periods of gradual evolution are possible, however, only if the structure of the organism makes it possible for random variations to try out a wide range of new traits. In the animals mentioned above, what makes it possible to accumulate traits in that way is their multicellular structure. Each cell together with its behavior is at least one of the structural causes that is bundled together as parts of a multicellular animal going through a reproductive cycle, and what makes it possible to bundle such parts together is its way of coordinating their behavior as parts of an organism.
Random variations ultimately involve differences in genes, the most elementary structural cause with both essential kinds of structural effects in living objects. But as we shall see, genetic variations make it possible not only for cells to try out specializing in new functions, but also for multicellular organisms to try out new ways of arranging such cells. The latter aspect of random variations is possible because each multicellular animal is constructed by asexual division from a single, fertilized egg cell in a process called “embryological development.”
The kinds of random variations that can be tried out at any point determines the range of new causal connections that are possible at that point. And since they occur randomly, all but a few whose complexity borders on making them impossible would eventually be tried out in a finite period of time. Thus, random variations on existing multicellular structures are, in effect, continually “trying out” new, possible powers, as if they were feeling around for new conditions that it might be useful to control. Whenever a new trait happened to control a new relevant condition (or an old condition in better way), it would tend to be selected. Moreover, as multicellular organisms became more complex, new varieties would be added whenever some were able to tap a new source of energy or an old one in new ways, and each variety would become more powerful in its ecological niche.
The principle of gradual evolution.
The example of the evolution of amphibians suggests a basic principle about
gradual evolution. If evolution is by reproductive causation, then every
power that it is possible for such organisms to evolve will evolve as it becomes
possible. This may seem too progressive to be true, but consider the following.
(1) If organisms start out simple and uniform, all possible traits are likely to be tried out, because the range of random variations is not very large. And later on, when organisms are more complex and the range of possible random variations is greater, there will be many more varieties of organisms to try them all out.
This is to assume that a complex variety that acquires a new basic power which makes it better able to tap the usable energy claimed by another will tend to supplant the other. That may require violent storms or other disruptions.
(2) It is not necessary for a random variation to happen on the best way of controlling the relevant condition. Even a weak and unreliable way of controlling some new relevant condition would be selected, and as random variations on it were tried out, that structural cause would be shaped to control it as effectively as possible and to fit it together with other structural causes and their causal connections as harmoniously as possible.
(3) Nor is it plausible to suppose that there are possible powers that just happen not to be tried out, at least, not any basic ones, because organisms are not being forced to make do with whatever random variations are available at the time in order to adapt to externally caused changes in the environment. When the cause of natural selection is reproduction, they have all the time they need for random variations to “feel around” for new powers among alternative ways of controlling relevant conditions and make those traits maximally efficient.
Maximum holistic power. Natural perfection in organisms is maximum holistic power, or the maximum power of the organism to control all the relevant conditions using the fewest and simplest non-reproductive structural effects. That is the optimal part-whole relation in organisms. Let me explain these terms.
Power. What is maximized is power. Power is the capacity of structural causes using free energy to make things happen that would not otherwise happen. The power that is maximized in organisms is the power of its structural causes to do non-reproductive work.
Holistic. The relevant power is holistic, because it is the result of many structural causes working together in some way over the period of the reproductive cycle. Organisms are bundles of structural causes that are reproduced as a whole during the cycle, and each structural cause is responsible for a non-reproductive structural effect, or what is usually called a “trait.” Each such structural cause is a part of the organism that generates a structural global regularity in which free energy is used to make something happen in the world that would not otherwise normally happen, and with many different structural causes bundled together in the organism, their structural effects may occur at any time during the reproductive cycle (including some that occur during the whole cycle, such as the circulation of blood in multicellular animals). Thus, the relevant power of the organism is holistic, because it includes all the non-reproductive work its structural causes do during its whole reproductive cycle.
Relevant. The structural effects that are relevant to the maximum holistic power of the organism are those that control conditions that affect its reproduction. Though it is possible for structural causes to control many kinds of conditions in the world, not all such powers are relevant to the optimal part-whole relation in organisms. Powers are not relevant if they make no difference to whether or not the organism reproduces. In order to contribute to the maximum holistic power of the organism, structural causes must control some relevant condition. (However, this limitation on the powers that are relevant changes, as we shall see, with the evolution of rational animals.)
Maximum. Holistic power is maximum when the organism’s non-reproductive structural effects control as many of the conditions affecting its reproduction that it is possible for it to control.
The powers that are possible are those that are within the range of those tried out by random variations on evolving organisms as they are evolving, including both variations in kinds of structural causes and variations in how they are arranged in the organism (perhaps at several levels of part-whole complexity).
The whole is not just the sum of its parts, because the structural causes depend on one another in various ways to control relevant conditions. The whole is both spatial and temporal, because the structural causes are different parts of the complex material structure that goes through the reproductive cycle (that is, the organism), and they have their structural effects at various times during the cycle. For the power of the whole to be maximum, therefore, the structural causes must be of the right kinds and numbers and they must combined in the right ways. That means that all the relevant conditions must be controlled with the fewest and simplest structural effects, for there is a cost in generating any irreversible structural effect.
When holistic power is maximum, the organism does the most with the least. It is not just the greatest combined power to control relevant conditions, but such a maximum using the fewest and simplest structural causes. Such an optimal part-whole relation is like maximum efficiency, the goal in designing machines. But I will call it “maximum holistic power,” rather than maximum efficiency, because I do not want to suggest that there is some overall function that organisms are serving. The optimum for organisms involves controlling all relevant conditions that can be controlled by organisms of its kind, that is, maximum holistic power to control relevant conditions.
In sum, the natural perfection toward which the gradual evolution of organisms proceeds is their maximum holistic power to control relevant conditions.
Every relevant power evolves as it becomes possible. The powers are the effects of structural causes that are bundled together as the organism going through the reproductive cycle, which are usually identified as traits serving some function. Each kind of structural cause is shaped to make its effect as powerful as possible in controlling some relevant condition. Structural causes are added to the organisms, both new kinds or more of old kinds, as long as they control some new condition affecting reproduction or some old condition more effectively. And the structural causes are shaped so the their effects fit in with the other traits as harmoniously as possible.
Thus, assuming that organisms start out simple, uniform and barely able to complete cycles of reproduction at all, they become more complex, diverse, and powerful. Eventually, all the structural causes that could help control conditions that affect the reproduction of organisms tapping some source of energy will be bundled together harmoniously in a reproductive cycle. Such gradual evolutionary change must stop only when there are no changes within the range of possible random variations that can make them more powerful in controlling relevant conditions. Though this end may be approached asymptotically, evolution will be in the direction of bundles of structural causes whose power, taken together over the whole cycle, is maximum.
Functional explanation. Since the gradual evolution of organisms is in the direction of natural perfection, there is an ontological definition of “functional,” one that entails the validity of functional explanations. This implication of spatiomaterialism solves various philosophical problems about the nature of functions, which are discussed in Epistemological philosophy of causation. But functions must be mentioned here, because I will talk about them and use them to explain what evolves. And though traits of organisms are only one sort of thing that is functional, if evolution is due to reproductive causation, I will discuss how they are functional here, since they are the focus of attention in contemporary biology.
In other words, there are "ends" built into nature, much as the teleological view of nature has long supposed. But change does not occur for the sake of such ends because of final causation, as Aristotle believed. Instead, such changes are inevitable products of evolution. They are consequences of the reproductive cycle as an ontological cause of global regularities.
Functions as descriptions of traits. The functions of traits are the relevant conditions that they control. What is bundled together as an organism going through reproductive cycles are structural causes, and as organisms change gradually in the direction of natural perfection (for organisms of their kind), structural causes come to control conditions that affect their reproduction. Those relevant conditions are the functions served by the traits of the organism.
The function of a non-reproductive structural effect is to control some condition that affects the reproduction of the organism of which it is part, and being functional in that way is what it contributes to the natural perfection of the organism.
Thus, any of the objects, events or conditions that are involved in the structural cause bringing about its non-reproductive structural effect can be said to be functional and to have the function of controlling the relevant condition involved, including all the features of organisms that are identified as its traits. Thus, not only the heart, but also the beating of the heart, is functional, because it is part of the structural effect by which energy is distributed to all parts of the animal body.
To be functional is to be good. Since being functional entails controlling a relevant conditions, it contributes to the natural perfection of the organism, and that is the definition of “good.” Functionality is a form of goodness.
Functions as explanations of traits. Functions are not, however, merely a way of classifying traits by the relevant condition they control, for they are also causes of traits and can be used to explain them. The relevant conditions that must be controlled in order for the organism to be naturally perfect are in the cards, so to speak, because, as we have seen, every possible power inevitably becomes actual as it becomes possible. What it is possible to control depends on the range of structural causes that are tried out as random variations on the evolving organisms, and since those structural causes among them that promote the reproduction of the organism will eventually be naturally selected, the organism will inevitably acquire all possible structural causes that control some relevant condition. Thus, the organism acquires them because they are functional, and the function can be said to be the cause of the evolution of the trait that serves them. The function explains, therefore, the existence of the trait in the organism.
This causal connection seems puzzling, because the traits are also the efficient causes of the conditions they control, which are said to be their functions. The heart causes the circulation of the blood that is said to be its function. But there is a reverse causal connection in which the functions cause their traits. The function of circulating the blood is what causes the heart to evolve. What makes this true is that the former is a case of structural causes generating a structural global regularity, whereas the latter is a case of reproductive causation generating a reproductive global regularity, namely, the global regularity in which all possible powers necessarily become actual.
Since functionality is a form of goodness, it follows that the goodness of the trait can also be said to be what causes it to evolve. Among the possible traits, those that are good are naturally selected because they contribute to the maximum holistic power and, thus, the natural perfection of the organism.
“Survival and reproduction.” Finally, if evolution is by reproductive causation, it is misleading to say that traits "contribute to the organism’s survival and reproduction.” Though it may not be false to say that traits are selected for contributing to the organism’s survival and reproduction, that phrase makes it sound as though traits, and even survival, are merely means to the end of reproduction. But almost the opposite is true, if evolution is a global regularity caused ontologically by the reproductive cycle.
The end for which the traits are means is not reproduction, but controlling relevant conditions. The organism has the traits because they contribute to maximum holistic power in controlling conditions that affect its reproduction. The fact that natural selection is made by success in reproduction means that what is selected is the whole bundle of causal connections, rather than some particular trait. And as we have seen, since natural selection is also caused by reproduction, the bundle is selected for its power to control all the conditions that affect its reproduction over a whole reproductive cycle. Any particular trait is included only because it controls some relevant condition that would not otherwise be controlled.
Non-reproductive structural effects have functions, as we have seen, because they control relevant conditions. The relevant condition they control is their function. Though reproduction is also a structural effect, it has no function. Reproduction is merely the cause of evolutionary change in the direction of natural perfection, including the maximum holistic power of organisms. Thus, reproduction determines which conditions it is relevant to control. But that does not make reproduction the ultimate function of non-reproductive work. Evolution is not change in the direction of reproduction, but in the direction of the maximum holistic power of organisms, or their natural perfection.
In other words, what is good for the organism is not reproduction, but controlling conditions that affect reproduction (though the latter may include some that are more closely related to reproduction, such as a mating or caring from offspring in the case of animals). When the organism controls the relevant conditions, reproduction generally takes care of itself, since it has been part of the reproductive cycle from the beginning.
![]() Gradual
evolution of the ecology. Implicit in the gradual evolution of
organisms is the gradual evolution of their combination in regions of space,
or the ecology. If organisms start off simple, uniform and barely able to
complete reproductive cycles, then reproductive causation will make them not
only more complex and powerful, but also more diverse. The usable energy in
any region of space is not only finite, but takes many different physical
forms, such as various kinds of photons, energy-rich molecules, and other
organisms. Thus, as these organisms specialize in tapping different sources
of energy (or the same sources of energy in different ways), they radiate
into all possible ecological niches, and evolutionary change in them—and
in whatever other organisms may remain and adapt to them—will be in the direction
of their maximum consumption of free energy, which is the natural perfection
of the ecology.
Gradual
evolution of the ecology. Implicit in the gradual evolution of
organisms is the gradual evolution of their combination in regions of space,
or the ecology. If organisms start off simple, uniform and barely able to
complete reproductive cycles, then reproductive causation will make them not
only more complex and powerful, but also more diverse. The usable energy in
any region of space is not only finite, but takes many different physical
forms, such as various kinds of photons, energy-rich molecules, and other
organisms. Thus, as these organisms specialize in tapping different sources
of energy (or the same sources of energy in different ways), they radiate
into all possible ecological niches, and evolutionary change in them—and
in whatever other organisms may remain and adapt to them—will be in the direction
of their maximum consumption of free energy, which is the natural perfection
of the ecology.
At the ecological level, there is a part-whole relation which can be optimal in the way that is appropriate in a spatiomaterial world like ours. The parts are the organisms, or bundles of structural causes going through reproductive cycles, and the whole is not only how the organisms are combined spatially in the region, but also how their reproductive cycles are combined in time over a period that is long enough to include all and any regular changes in the environment, such as seasons. The species of organisms that evolve during any radiation tend to mirror the sources of usable energy in the region. As each species becomes more efficient and approaches the maximum holistic power for organisms of its kind, reproductive causation is also making their combination in the region optimal, but in a different way from the organisms.
The effect of the whole combination of organisms that is being maximized is the consumption of free (or usable) energy to fuel cycles of reproduction. Any unused free energy is an ecological niche to be occupied, if traits being tried out by random variations on any existing organism enable it to tap the energy as fuel for its reproductive cycles, for that overcomes the scarcity due to population growth. Thus, all possible ways of tapping usable energy come to be included in the ecology. And organisms are combined in the most efficient quantities, since the finite amount of usable energy is what stops population growth and causes natural selection in each species. The maximum may be approached only asymptotically, but as reproductive causation maximizes the power of the organisms over their whole reproductive cycle, evolutionary change at the ecological level is in the direction of appropriating more and more the usable energy in the region to fuel reproductive cycles, making the ecology as a whole maximally powerful in the way that is appropriate to a spatiomaterial world like ours.
Evolution by reproductive causation is change in the direction of natural perfection for both the individual organisms and their ecology. Indeed, it is only because the organisms have maximum holistic power that the ecology does the most with the least.
Natural perfection at the level of the ecology is using as much of the available free energy as possible to fuel the reproductive cycles of maximally efficient organisms of many varieties. But it does so by combining the simplest and fewest organisms—simplest because maximum holistic power at the level of individual organisms is using as little usable energy as possible to exert the greatest power to control all the conditions that affect its reproduction, and fewest because there are no more organisms than the supply of usable energy will support. Thus, these two forms of natural perfection are opposite sides of the same optimum: using the most energy to maximum effect. In other words, gradual evolution is change in the direction of a “compound” natural perfection, because the optimum for the ecology is a whole made up of parts that are, themselves, optimal as organisms.
This kind of perfection is rightly called "natural," because it makes the most of the basic nature of what exists in a spatiomaterial world like ours. The only possible way of making the most of space and matter in time is using free energy to control what happens in the world. At the end of gradual evolution, not only does the ecology consume as much of the free energy available to control conditions in the world as possible, but also all the conditions in the world that can be controlled by them are controlled (because the organisms are as powerful as possible for organisms of their kind and they use as little free energy as required for that maximum).
The natural perfection of the ecology has the same implications for organisms that its member the natural perfection of the organism has for its traits. Just as the organism's traits are functional because they control some relevant condition, so organisms in the ecology are functional because they consume some of the available free energy. And since to be functional is to be good, organisms are also good because they consume the free energy in some form or way. That is what the organism is good for, as far as the natural perfection of ecology is concerned.
The analogy among kinds of natural perfection suggest a further step. There is, as I suggested earlier, another kind of natural perfection about gradual evolution, namely, the way in which reproductive causation brings about the natural perfection of the organism and the ecology. Each phase of the process of gradual change in that direction makes a necessary contribution to their existence at the end, and since that is also an optimal part-whole relation, each phase can also be said to have such a function and to be good for contributing to the perfection of the whole in that way.
Finally, if natural selection is caused by reproduction, change in the direction of this compound natural perfection is inevitable, for it does not depend on externally caused changes in the environment. To be sure, evolutionary change itself may be seen as a change in the environment. As organisms become more powerful, traits that random variations have been trying out all along but were neutral or even harmful may suddenly become useful in promoting reproduction because of how they work together with other traits that have been acquired in the meantime. Or traits that first evolved to control one condition may come to control others. And evolutionary change in one species may change the effects of the traits of other species, requiring them to adapt or become extinct. But if such changes in the effects of traits are environmental changes, they are not caused externally. They come from the increasing power of the evolving organisms in whatever environment they all inhabit and, thus, are internal to evolution by reproductive causation, like scarcity due to population growth.
![]()